 2010, Vol. 46
2010, Vol. 46文章信息
- 杨高鹏, 段立清, 宫玉艳, 杨帆, 孙彦宏
- Yang Gaopeng, Duan Liqing, Gong Yuyan, Yang Fan, Sun Yanhong
- 荧光增白剂、寄主植物、LdNPV地理品系对舞毒蛾幼虫酚氧化酶的影响
- Effect of a Fluorescent Brightener, Host Plants and Geographic Strains of LdNPV on the Phenoloxidase Activity of Gypsy Moth Larvae
- 林业科学, 2010, 46(9): 105-109.
- Scientia Silvae Sinicae, 2010, 46(9): 105-109.
-
文章历史
- 收稿日期:2009-03-24
- 修回日期:2009-06-10
-
作者相关文章



舞毒蛾核型多角体病毒(Lymantria dispar nuclear polyhedrosis virus, LdNPV)是影响舞毒蛾种群动态的主要因子之一,在美国、加拿大、俄罗斯等地的防治中均取得了良好的效果; 但病毒杀虫剂由于杀虫范围窄、作用速度较慢等原因,生产应用不广泛,目前仅占整个农药市场的0.2%(郭慧芳等,2003)。
Shapiro(1992)研究发现荧光增白剂不仅具有保护昆虫病毒免受紫外线破坏的作用,而且还可提高病毒本身的毒力。此后,相继发现荧光增白剂在舞毒蛾、秋粘虫(Spodoptera frugiperda)、棉铃虫(Helicoverpa armigera)、大豆夜蛾(Pseudoplusia includens)、芹菜夜蛾(Syngrapha falcifera)、云杉卷叶蛾(Choristoneura fumiferana)、小地老虎(Agrotis ypsilon)和甜菜夜蛾(Spodoptera exigua)等多种害虫中对核型多角体病毒的增效作用(郭慧芳等,2003)。Tinopal LPW因其纯度高并对多种昆虫病毒与昆虫作用体系表现有极其显著和稳定的增强作用,被视为研究荧光增白剂(二苯乙烯类)对昆虫病毒增强作用的标准样品,是目前能增强昆虫病毒种类最多的荧光增白剂(严东辉等,2003)。关于荧光增白剂增强昆虫病毒毒力的作用机制,目前还很难从荧光增白剂本身的结构特点和荧光增白剂与昆虫病毒二者之间的关系来探索。研究靶标集中在昆虫中肠上,荧光增白剂究竟是作用于中肠细胞还是围食膜,除此之外,有无其他靶标,这些都还需要进一步研究。
昆虫体内存在着丰富的酶类,其中酚氧化酶(phenoloxidase, PO)又叫酪氨酸酶是昆虫体内最重要的免疫因子之一,它主要以无活性的酶原形式存在于昆虫的血淋巴、中肠和表皮等组织中。在血淋巴中能导致昆虫抗菌醌类物质的形成及寄主血细胞的黑化(袁勤生,2001)。昆虫主要通过产生黑色素杀死入侵寄生物,酚氧化酶参与黑色素的形成,在昆虫体液免疫中起着重要作用(时超美,2000)。另外酚氧化酶还参与醌鞣化作用,促进昆虫表皮硬化,在昆虫发育过程中扮演重要角色,同时酚氧化酶还被认为是昆虫表皮鞣质化和伤口愈合,以及抵御病原生物起重要作用的酶(Ashida et al., 1984; Sugumaran, 1998; Taukamoto et al., 1992; Theopold et al., 2004)。鉴于酚氧化酶在昆虫防御反应中的关键作用,对其活力的测定是研究昆虫免疫反应的基础(尹丽红等,2001)。为探索昆虫病毒的致病及荧光增白剂的增效机制,选择舞毒蛾5龄幼虫作为试验材料,研究病毒品系、荧光增白剂、寄主植物对舞毒蛾酚氧化酶比活力的影响。
1 材料与方法 1.1 材料、试剂及仪器设备舞毒蛾卵块采自内蒙古自治区通辽市,-4 ℃冷藏备用。华北落叶松(Larix principis-rupprechtii)、山杏(Prunus mandshurica)、青杨(Populus cathayana)均采自内蒙古农业大学科技园区。LdNPV病毒品系由加拿大太平洋森林中心(PFC)提供,分别为北美品系(LdNPV-D)、中国黑龙江品系(LdNPV-H)和日本品系(LdNPV-J)。
荧光增白剂Tinopal LPW(SIGMA公司产品)、苯基硫脲(N-phenylthiourea)(SIGMA公司产品)、脯氨酸(L-proline)(日本五洲生物公司)、柠檬酸(citric acid)、柠檬酸钠(trisodium citeate dihydrate)、邻苯二酚(O-dihydroxybenzne)、偏磷酸(metaphosphoric acid)。高速冷冻离心机(Hettich Zentrifugen GmbH & Co. KG)、TU-1810紫外-可见分光光度仪(北京普析通用仪器有限公司产品)、电子天平(北京赛多利斯天平有限公司产品)、移液枪(法国GILON公司)。
1.2 舞毒蛾的饲养与处理舞毒蛾卵块去附毛,1% “84”消毒液中浸泡10 s,蒸馏水冲洗3次后置于气候培养箱(t=25 ℃, RH=60%, L:D=16:8)内孵化。孵化后分别用以上3种植物饲喂至5龄,选择同24 h内进入5龄的大小、体质量相近的虫体饥饿24 h供试。
设6个处理:1×106 OBs·mL-1 H品系及其与1%Tinopal LPW的混合液; 1×106 OBs·mL-1的D品系及其与1%Tinopal LPW的混合液; 1×106 OBs·mL-1的J品系及其与1%Tinopal LPW的混合液。以1%Tinopal LPW和蒸馏水作对照。
将山杏、青杨叶剪成面积为1 cm2方形,华北落叶松叶簇剪成直径为1 cm一簇,每个叶片(簇)接毒30 μL处理液,晾干。每种处理需150个叶片(簇),每种寄主植物8个处理,分别需1 200个叶片(簇),共需3 600个叶片(簇)。将处理的叶片(簇)放入养虫杯,每杯1叶片(簇),接入饥饿24 h后的舞毒蛾5龄幼虫1头。取食6 h后,挑取食完全部叶(簇)的幼虫接以上3种寄主植物上继续饲养。分别于12, 24, 48, 72, 96 h取幼虫,用酒精擦拭消毒后放在冰盘中,自腹足处用1 mL注射器取血淋巴,将血淋巴收集到1 mL离心管中。为防止氧化,离心管中先加入微量苯基硫脲。
1.3 酚氧化酶活性测定将上述血淋巴在4 ℃ 3 000 r·min-1离心5 min,取上清液作为酶液。参照李季生等(2006)的方法,用TU-1810紫外-可见分光光度仪460 nm波长处测定OD值,每处理重复3次。
1.4 蛋白含量测定参照Bradford(1976)的方法,用考马斯亮蓝染色,以牛血清白蛋白(BSA)作为标准蛋白。
1.5 数据处理采用SPSS 13.0数据处理软件进行方差分析,并用Duncan’s检验差异显著性。
2 结果与分析 2.1 寄主植物对舞毒蛾5龄幼虫PO活力的影响取食青杨﹑华北落叶松和山杏的舞毒蛾5龄幼虫血淋巴中PO比活力变化见图 1。

|
图 1 取食不同寄主植物对舞毒蛾幼虫PO比活力的影响 Figure 1 The effect of different host plants on PO specific activity of gypsy moth larvae 图中相同字母表示差异不显著,不同字母表示差异显著; P=0.05(n=30);括号内字母表示相同寄主植物不同时间段下舞毒蛾幼虫PO的差异; 不带括号字母表示同一时间段不同寄主植物下舞毒蛾幼虫PO的差异。The same letter show the no obvious difference of treatments and the different letters show the obvious difference of treatments; P=0.05(n=30);Letters in the bracket show the significance of PO activity of gypsy moth fed on the same host plant at different time; letters outside of bracket show the significance of PO activity of gypsy moth fed on different host plants at the same time. |
由图 1可知,舞毒蛾5龄幼虫血淋巴中PO比活力变化呈抛物线型,取食青杨和华北落叶松的幼虫PO比活力在48 h最大,取食山杏在72 h最大。寄主植物对舞毒蛾幼虫的PO比活力影响差异显著。
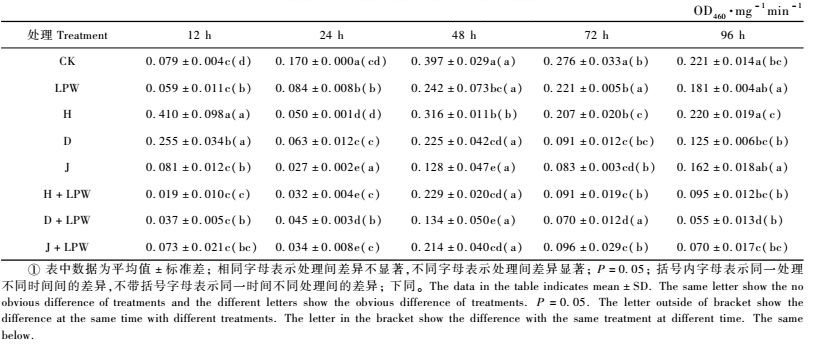
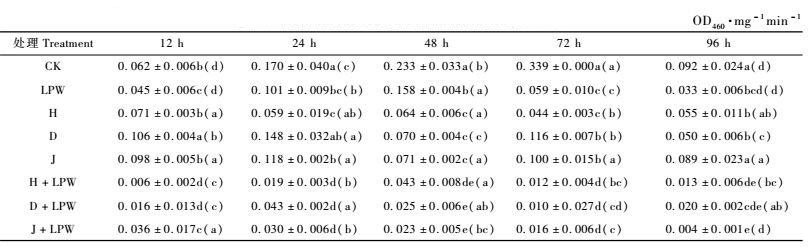
2.2 同一寄主不同处理对舞毒蛾幼虫PO的影响Tinopal LPW分别与LdNPV的3个品系混合,感染取食青杨﹑落叶松和山杏的舞毒蛾5龄幼虫,同一寄主植物、不同处理对舞毒蛾5龄幼虫各阶段血淋巴PO比活力的变化见表 1, 2, 3。
|
|
|
|

|
|
由表 1可知,以青杨为寄主的舞毒蛾5龄幼虫在12 h时H处理和D处理的PO活性均显著高于清水及LPW对照,其他处理与对照均无显著差异。24 h时各处理及LPW对照的PO值均显著低于清水对照,各处理也均低于LPW对照。48 h时各处理及LPW对照的PO活性均显著低于清水对照。72 h时各处理及LPW均显著低于清水对照,除H处理外,各处理的PO活性均显著低于LPW处理。96 h时,除H处理和J处理外,其他处理的PO活性均显著低于清水对照,D品系和J品系的Tinopal LPW混合液对PO活力的抑制显著低于LPW对照。H, D和J处理96 h的PO活性均较72 h时活性增加,PO活性变化趋势与对照不同。
由表 2可知,以华北落叶松为寄主植物的舞毒蛾5龄幼虫在12 h时,除J品系外,其他2个品系处理与清水对照无显著差异,而3个品系处理的PO活性均显著高于LPW对照。3个病毒品系的Tinopal LPW混合液处理的PO活性均显著低于清水对照,H品系的Tinopal LPW混合液处理的PO活性显著低于LPW对照。24 h时除D处理外, 其他处理的PO活性均显著低于清水及LPW对照。48 h时所有处理的PO活性均显著低于清水对照,3个病毒处理与LPW对照无显著差异,3个病毒品系的Tinopal LPW混合液处理均显著低于LPW对照。72 h时所有处理的PO活性均显著低于清水对照,H品系和D品系的LPW混合液处理的PO活性显著低于LPW对照。96 h时所有处理的PO活性均显著低于清水对照,D处理、H品系和D品系的Tinopal LPW混合液对PO活力的抑制显著低于LPW对照。
由表 3可知,以山杏为寄主植物的舞毒蛾5龄幼虫在12 h时,D处理的PO活性显著高于清水对照,3个病毒品系处理的PO活性均显著高于LPW对照,H品系和D品系的Tinopal LPW混合液对PO活力的抑制显著低于清水及LPW对照,而J品系的Tinopal LPW混合液处理PO活性只与清水对照有差异。24 h时除D处理外, 其他处理的PO活性均显著低于清水对照,3个病毒品系的Tinopal LPW混合液使PO活性显著低于LPW对照。48 h时所有处理的PO活性均显著低于清水和LPW处理。72 h时所有处理的PO活性均显著低于清水对照,96 h时除J品系外,其他处理均显著低于清水对照,只有J品系的Tinopal LPW混合液处理的PO活性与LPW差异显著。
不论取食何种植物,荧光增白剂LPW处理的PO活性均显著低于清水对照,说明Tinopal LPW对舞毒蛾5龄幼虫血淋巴PO有抑制作用。
3 结论与讨论舞毒蛾5龄幼虫血淋巴中PO比活力分布呈抛物线型,同其他昆虫的研究结果类似,如亚洲玉米螟(Ostrinia furnacalis)4龄幼虫后期血淋巴中PO活性最高,蜕皮后迅速下降,然后随5龄幼虫的生长,PO活性逐渐升高,到第7天,其活性又达最高值(程振衡等,1990); 棉铃虫被中华卵索线虫(Ovomermis sinensis)寄生后的第1天PO活性比对照组高,随后比对照低,而对照组酚氧化酶的活性在第5天达最高(李志强等,2006)。寄主植物对舞毒蛾幼虫的PO比活力影响差异显著,这可能与寄主植物的PO活性、营养物质或次生物质有关,有待进一步深入研究。酚氧化酶是昆虫防御体系中重要的酶和免疫因子,一旦有异物入侵时,酶活性及结构会发生变化。食入纯病毒的舞毒蛾幼虫,在12 h时PO活性均高于清水对照,说明病毒的侵入刺激了舞毒蛾幼虫的免疫系统,激活PO,产生抵抗。随着病毒在体内的不断复制扩增逐渐抑制了舞毒蛾幼虫的免疫系统,使其PO降低。
Sheppard等(1994)认为Tinopal LPW对LdMNPV的增效作用与中肠pH值下降有关。但Shapiro(2000)认为中肠pH值下降是病毒感染中肠细胞的结果。Washburn等(1998)则认为中肠pH值下降是因为荧光增白剂阻止了被病毒感染中肠上皮细胞的脱落,是增效的主要原因。而Wang等(2000)对围食膜的研究表明,荧光增白剂Calcoflour通过竞争性地结合在几丁质上,阻止蛋白质的结合而破坏围食膜,这是增效的原因。张丽红等(2004)研究发现荧光增白剂28增强了病毒粒子与中肠上皮细胞的融合,认为这是增效的原因。食入Tinopal LPW,使舞毒蛾幼虫的PO活性降低,说明Tinopal LPW对PO有抑制作用,降低其对病毒侵染的抵抗能力,增加了病毒侵染几率,起到增效作用。
探索荧光增白剂、寄主植物、LdNPV地理品系对舞毒蛾幼虫血淋巴PO的影响, 从理论上较全面地了解寄主植物、病毒品系及其增效剂的相互作用, 对危害不同林木的舞毒蛾进行防治时,可为病毒品系的选择、增效剂的利用提供理论依据。
李季生, 夏爱华, 牟志美, 等. 2006. 蝇蛆寄生对家蚕血细胞和酚氧化酶活性的影响[J]. 蚕业科学, 32(2): 268-271. |
李志强, 王茂先, 陈国生. 2006. 棉铃虫感染中华卵索线虫后血淋巴酚氧化酶活性的变化及其分离纯化[J]. 昆虫学报, 49(5): 878-881. |
郭慧芳, 方继朝, 韩召军. 2003. 昆虫病毒增效剂研究进展[J]. 昆虫学报, 46(6): 766-772. |
程振衡, 梁子才. 1990. 亚洲玉米螟血淋巴中酚氧化酶的研究[J]. 昆虫学报, 33(4): 424-429. |
时超美. 2000. 昆虫酚氧化酶原活化及其在免疫中的作用[J]. 昆虫知识, 37(5): 310-311. DOI:10.7679/j.issn.2095-1353.2000.089 |
严东辉, 陈昌杰, ShapiroM. 2003. 提高昆虫病毒杀虫效果的荧光增白剂的研究[J]. 林业科学, 39(1): 153-159. DOI:10.11707/j.1001-7488.20030125 |
尹丽红, 王琛柱, 钦俊德. 2001. 棉铃虫血淋巴酚氧化酶活性的微量测定[J]. 昆虫知识, 38(2): 119-122. |
袁勤生. 2001. 现代酶学[M]. 上海: 华东理工大学出版社: 3-18.
|
张丽红, 彭建新. 2004. 光增白剂对杆状病毒的增效作用及其作用方式研究. 华中师范大学硕士学位论文.
|
Ashida M, Soderhall K. 1984. The prophenoloxidase activating system in crayfish[J]. Comparative Biochemistry and Physiology, 77B(1): 21-26. |
Bradford M M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of prote in-dye binding[J]. Analytical Biochemistry, 72(1-2): 248-254. DOI:10.1016/0003-2697(76)90527-3 |
Shapiro M. 1992. Use of optical brighteners as radiation protectants for gypsy moth (Lepidoptera: Lymantriidae) nuclear polyhedrosis virus[J]. Journal of Economic Entomology, 85(5): 1682-1686. DOI:10.1093/jee/85.5.1682 |
Shapiro M. 2000. Effect of two granulosis viruses on the activity of the gypsy moth (Lepidoptera: Lymantiidae) nuclear polyhedrosis virus[J]. Journal of Economic Entomology, 93(6): 1633-1637. DOI:10.1603/0022-0493-93.6.1633 |
Sheppard C A, Shapiro M. 1994. Physiological and nutritional effects of a fluorescent brightener on nuclear polyhedrosis virus-infected Lymantria dispar (L.) larvae (Lepidoptera: Lymantriidae)[J]. Biological Control, 4(4): 404-411. DOI:10.1006/bcon.1994.1051 |
Sugumaran M. 1998. Unified mechanism for sclerotization of insect cuticle[J]. Advances in Insect Physiology, 27: 229-334. DOI:10.1016/S0065-2806(08)60014-4 |
Tsukamoto T, Lchimaru Y, Kanegae N, et al. 1992. Identification and isolation of endogenous insect prophenoloxidase inhibitions[J]. Biological Control, 184(1): 86-92. |
Theopold U, Schmidt O, Dushay M S, et al. 2004. Coagulation in arthropods: Defence, wound closure and healing[J]. Trends in Immunology, 25(6): 289-294. DOI:10.1016/j.it.2004.03.004 |
Washburn J O, Kirkpatrick B A, Haas-Stapleton E, et al. 1998. Evidence that the stilbene-derived optical brightener M2R enhances Autographa califoica nucleopolyhedrovirus infection of Trichoplusiani and Heliothis virescens by preventing sloughing of infected midgut epithelial cells[J]. Biological Control, 11(1): 58-69. DOI:10.1006/bcon.1997.0572 |
Wang P, Granados R R. 2000. Calcofluor disrupts the midgut defense system in insects[J]. Insect Biochemistry and Molecular Biology, 30(2): 135-143. DOI:10.1016/S0965-1748(99)00108-3 |