 2010, Vol. 46
2010, Vol. 46文章信息
- 高宝嘉, 杜娟, 高素红, 刘军侠
- Gao Baojia, Du Juan, Gao Suhong, Liu Junxia
- 美国白蛾种群的遗传多样性与遗传分化
- Genetic Diversity and Differentiations of Fall Webworm (Hyphantria cunea) Populations
- 林业科学, 2010, 46(8): 120-124.
- Scientia Silvae Sinicae, 2010, 46(8): 120-124.
-
文章历史
- 收稿日期:2009-04-24
- 修回日期:2009-05-18
-
作者相关文章


2. 河北北方学院 张家口 075000;
3. 廊坊师范学校 廊坊 065000;
4. 河北科技师范学院 秦皇岛 066600
2. North University of Hebei Zhangjiakou 075000;
3. Langfang Teachers College Langfang 065000;
4. Hebei Normal University of Scicence & Technology Qinhuangdao 066600
近年来,生态和昆虫学家就昆虫种群遗传多样性与种群分化进行了较多研究。学者们应用多种分子标记技术,探讨了昆虫的遗传多样性变化和种群的遗传分化。研究发现,昆虫种群的遗传多样性大多较高,群体间的遗传分化明显; 昆虫种群的遗传分化受环境因素的影响而且与种群的适应性有关。刘宁(2003)、薛进等(2007)的研究认为,由于地理位置的差异而阻隔了昆虫种群之间的基因交流,进而导致了种群遗传分化。乔传令等(2000)的研究发现不同用药情况将会导致不同的抗性水平,在一定程度上导致昆虫不同地理种群的特异性。袁一杨等(2008)、张爱兵等(2004)的研究还表明,不同生境、不同立地条件以及空间上的隔离都能对昆虫种群遗传多样性产生较大的影响。
美国白蛾(Hyphantria cunea)作为入侵性极强的有害昆虫,分布范围较广,生境条件多样,寄主植物种类繁多,扩散速度快,给国内的各个发生区造成了严重的经济损失(杨忠岐等,2007; 季荣等,2003)。美国白蛾入侵中国的近30年中,我国学者对它的适应性、性信息素、对寄主的选择、天敌、预测预报和防治方法等方面作了广泛研究(季荣等,2003; 魏建荣,2003; 杨忠岐等,2007; 朱丽红等,1998)。然而,关于该入侵害虫的遗传多样性和遗传变异规律尚缺少深入研究。本文采用微卫星分子标记技术对5个美国白蛾的遗传多样性与遗传分化进行分析,旨在揭示美国白蛾种群的遗传多样性水平、种群间和种群内的遗传分化程度和基因流,并据此分析其遗传结构的形成原因及其影响因素,从而预测美国白蛾的潜在入侵趋势,为有效地防治入侵美国白蛾提供基础理论依据。
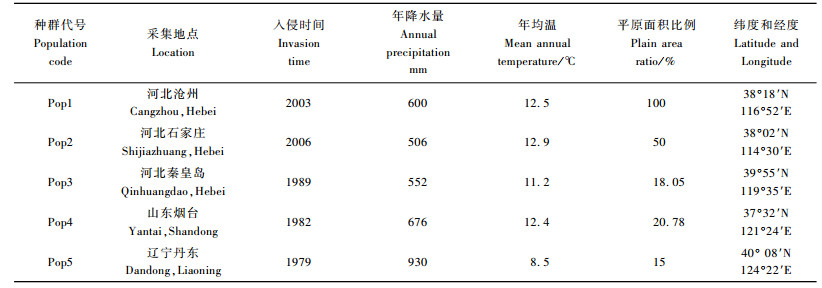
1 材料与方法 1.1 材料采集美国白蛾5个种群分别采自河北省沧州、石家庄、秦皇岛、辽宁省丹东、山东省烟台(表 1,数据来源于当地的林业部门),在采集地随机采集虫蛹带回室内,成虫羽化后立即放入-20 ℃冰箱保存。
|
|
取美国白蛾的胸腹部肌肉,研磨磨碎后加入适量消化液,用酚/氯仿法抽提,用0.8%琼脂糖凝胶电泳检测DNA的纯度,-20 ℃保存备用。
1.3 PCR扩增与电泳检测引物筛选:对购自上海生工公司的52对引物和北京海泰克公司的8对引物进行了筛选(Ji et al., 2005;Chiharu et al., 2005; 沈利,2004; 李竟等,2005),从中选取扩增条带清晰,重复性较好的10对微卫星引物(表 2)进行扩增,扩增反应在美国ABI公司生产的2720型PCR仪上进行。
|
|

扩增反应条件:25 μL反应体系含0.4 mmol·L-1 dNTP,2.5 mmol·L-1 MgCI2,1 U Taq聚合酶,0.32 μmol·L-1引物,30~60 ng DNA。
扩增程序:94 ℃预变性4 min; 94 ℃变性20 s,退火30 s,72 ℃延伸15 s,35个循环; 最后72 ℃延伸2 min。
电泳检测:产物用8%非变性聚丙烯酰胺凝胶电泳,电泳缓冲液为1×TBE,用海泰克公司生产的100 bp Ladder Marker,照相保存。
1.4 数据统计与分析采用人工读带法,根据条带的迁移率和有无记录二元数据,有带记1,无带记0。排除模糊不清的带和无法准确标识的带。用POPGENE32计算群体遗传多样性和遗传分化等指数,并用UPGMA进行聚类分析。采用主成分分析(principal component analysis,PCA)技术可反映出种之间的相似性及有关性状间的更多信息。
2 结果与分析 2.1 美国白蛾SSR-PCR的扩增结果从60对引物中选取扩增条带清晰,重复性较好的10对微卫星引物,对300个个体的DNA样品进行PCR扩增,其序列见表 2,各对引物的退火温度是经过摸索而得到的。引物4对5个种群的扩增结果的杂合度最高,引物10的杂合度最低(表 2)。其中引物7对美国白蛾5个种群的25个个体的扩增结果如图 1所示。

|
图 1 引物7对5个美国白蛾种群的SSR扩增结果 Figure 1 SSR profile of 5 populations amplified by primer SSR7 1-5:种群5 Pop5;6-10:种群4 Pop4;11-15:种群3 Pop3;16-20:种群2 Pop2;21-25:种群1 Pop1; M:分子量标准Molecular marker(100 bp ladder). |
美国白蛾300个个体共检测到60个多态位点,总多态位点百分率为98.36%,不同种群中检测到的位点数及多态位点百分率不同(表 3)。10条引物对沧州种群检测出的位点数和多态位点百分率最多,分别为47和77.05%;石家庄种群和丹东种群的位点数和多态位点百分率最少,分别为43和70.49%。5个种群的多态位点百分率明显低于总的多态位点百分率。美国白蛾5个种群多态位点百分率的大小顺序为:沧州种群、烟台种群、秦皇岛种群、丹东种群、石家庄种群。
|
|
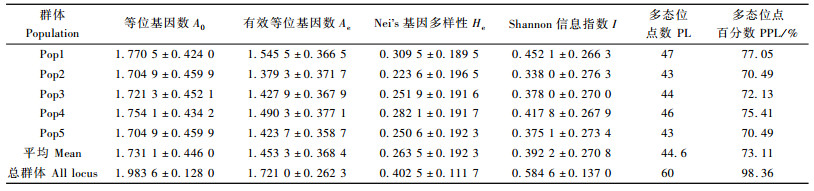
5个种群Shannon信息指数在0.338 0~0.452 1之间,平均值为0.392 2。沧州种群的Shannon信息指数最高,石家庄种群最低。5个种群Nei's基因多样性(He)变化范围在0.223 6~0.309 5之间,平均值为0.263 5。沧州种群的基因多样性最高,石家庄种群最低,这与Shannon信息指数估计的遗传多样性结果是一致的。
2.3 美国白蛾群体间的遗传分化美国白蛾群体总的基因多样性(Ht)为40.16%,种群内的基因多样性(Hs)为26.35%;美国白蛾群体的遗传分化系数(Gst):34.38%的变异存在于种群间,65.62%的变异存在于种群内; 美国白蛾从Gst估计的基因流Hm为0.954 2。
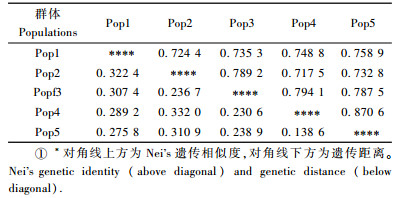
2.4 美国白蛾的遗传距离及聚类分析遗传距离是研究物种遗传多样性的基础,它反映了研究群体的系统进化,用以描述群体的遗传结构及种群间的差异,说明每个种群间彼此关系的远近。为确定美国白蛾5个不同入侵定殖区种群之间的遗传关系,对Nei's无偏遗传距离D进行了计算(表 4)。各种群遗传距离在0.138 6(丹东种群-烟台种群)~0.322 4(沧州种群-石家庄种群)之间。利用UPGMA方法对美国白蛾在中国不同入侵定殖后的种群进行聚类分析,分化时间最早的丹东种群和烟台种群最先聚为一支,然后依次与秦皇岛种群、沧州种群聚为一类,最后与分化时间最晚的石家庄种群聚类。由此可见,石家庄种群已经明显的独具一支,分化时间早的种群与分化时间晚的种群已产生明显的遗传差异,所以分化时间的早晚是影响种群分化程度的主要因子。
|
|

|
图 2 美国白蛾5个不同种群UPGMA聚类图 Figure 2 Phylogenetic relationship of 5 populations of fall webworm based on Nei's distance of SSR markers and clustered using UPGMA |
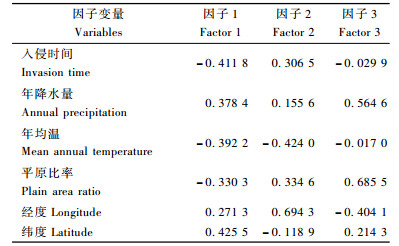
美国白蛾5个种群的寄主、防治强度(每年的用药次数,用药量,人为防治力度、方法,生物防治方法)大体相同,不同之处在于入侵时间、年降水量、年均温、平原比率、经纬度。将各种群不同的影响因子输入DPS V7.05软件做主成分分析,结果见表 5。以主成分特征值累计百分率>85%为标准,筛选出3个影响美国白蛾种群遗传多样性的主成分因子。在影响因子1中,纬度(0.425 5)得分最高,是主要影响因素; 在因子2中,经度对种群遗传多样性影响程度最高(0.694 3);在因子3中,平原比率得分高于其他生态因子(0.685 5),不同的平原比率意味着该地区美国白蛾寄主的多少,是美国白蛾在该地区扩散蔓延的范围大小。因此,经度和平原比率对美国白蛾种群的遗传多样性的影响程度最大,其次年降水量、纬度、入侵时间也均具有重要影响。
|
|
入侵我国的美国白蛾具有较为丰富的遗传多样性,通常认为,遗传多样性的高低反映了物种对外界环境适应能力的强弱,遗多样性丰富的外来生物适应性往往会很强,防治难度也较大。美国白蛾较为丰富的遗传多样性可能是其成功入侵定殖的因素之一。美国白蛾5个种群的遗传多样性统计表明,美国白蛾沧州种群Nei's的基因多样性最高(0.309 5),石家庄种群最低(0.223 6),该结论证明美国白蛾种群在中国不同入侵定殖区内遗传多样性存在较大差异。
美国白蛾喜潮湿的环境,多分布在我国沿海地区,多发生在城市、乡镇、居民区、公园等处的园林绿化树木和行道树以及果园等处(季荣等,2003)。通过对各种群不同的影响因子做主成分分析,结果表明经度与平原比率对美国白蛾种群的遗传多样性影响最大,其次年降水量、纬度、入侵时间也均具有重要影响,其原因主要与美国白蛾自身的生活习性有关。沧州地区是个沿海地区,经度偏高,平原比率为100%,从整个地区的特点看,无论是生境条件、立地条件都适合美国白蛾的生存与繁衍,笔者认为,这些特点是造成沧州种群遗传多态性高的主要原因。入侵物种常具有快速进化的能力,如通过遗传漂变、建立者种群的快速进化(Lee, 2002); 入侵种群的种内或种间杂交可能产生新的基因型,对新环境的适应等(Mooney et al., 2001; Sakai et al., 2001)。石家庄种群多态性低的原因主要是该地区的经度较其他4个地区偏小,气候偏干旱,西部是太行山脉,这些不适合美国白蛾的生存。另外,入侵该地区的时间最晚,也对种群多态性低起一定的作用。
另外,秦皇岛、烟台、丹东种群的遗传多样性均低于沧州种群并且高于石家庄种群,其原因主要是这3个地区都是丘陵山地地区,平原面积比例均小于沧州地区,都位于沿海,平均湿度都高于石家庄地区,遗传距离与经纬度和物候有着密切的关系。
3.2 美国白蛾遗传分化与影响因素影响自然群体遗传分化的原因有多个,主要包括基因流、突变和各种因素的选择作用(Taylor et al., 1990)等。不同环境条件引起的隔离是导致遗传分化的主要原因; 经纬度、温度、湿度等方面的差异都会导致群体间的遗传组成发生差异(李进等,1998); 顾少华等(1992)研究指出,地理位置越远,则基因频率的差异和遗传距离越大,而地理位置越近,则基因频率的差异和遗传距离越小。同样入侵物种的遗传变异可用来确定入侵物种的地理起源,也可确定是否是单个或多个种群起源。
美国白蛾种群内遗传分化系数远离于种群间,这说明美国白蛾的种群内遗传变异很大。高宝嘉等(2007)发现不同地域的油松毛虫遗传距离与地理距离有一定的相关性。本研究认为入侵物种的经纬度、平原比率、年降水量、入侵时间、入侵来源是导致种群变异的主要因素。美国白蛾种群入侵丹东、烟台地区的时间早且间隔较短,这2个地区是沿海地区,环境条件、地形地貌比较接近,地理隔离较小,因此这2个地区美国白蛾种群的遗传分化最小。1989年美国白蛾入侵秦皇岛地区,从当时的入侵来源看,该地区的美国白蛾种群是从丹东种群扩散而来的,其地区同样是沿海地区,与丹东地区的环境条件地形地貌相差不大,只是比丹东种群分化的时间较晚,因此分化程度仅次于丹东种群; 沧州种群是秦皇岛种群于2003年传入的,沧州种群比秦皇岛种群分化的更晚,因此分化程度次于秦皇岛种群; 石家庄种群的入侵来源是由丹东—秦皇岛—沧州一带传入的,分化时间较沧州种群更晚,因此分化程度也就更大。石家庄种群与烟台种群分化时间的早晚上有很大差异,与烟台种群的亲缘关系相比,石家庄种群与丹东种群的亲缘关系更近,因此,石家庄种群与烟台种群的分化程度最大。
此外,入侵物种遗传多样性及遗传分化可用于指导生物控制媒介的筛选,可以确定生物防治的敏感种群(Meekins et al., 2001; Nissen et al., 1995)。不同入侵地区的美国白蛾种群可能具有不同的起源,因此对美国白蛾的防治要多个地区同时进行可以提高生物防治的效率。另外,具有较高遗传变异的入侵物种的种群与生物控制媒介之间易发生抗性进化,导致防治难度增加(Sakai et al., 2001)。因此,要针对其遗传多样性与遗传变异进行深入探讨,以达到对美国白蛾有效的生物防治。
高宝嘉, 高立杰, 侯建华, 等. 2007. 三种松毛虫不同地理种群遗传多样性[J]. 生态学报, 28(2): 842-848. |
顾少华, 孔原, 朱定良, 等. 1992. 华东地区黑果蝇自然群体同工酶遗传多态的研究[J]. 遗传学报, 19(3): 228-235. |
季荣, 谢宝瑜, 李欣海, 等. 2003. 外来入侵种——美国自蛾的研究进展[J]. 昆虫知识, 40(1): 13-18. DOI:10.7679/j.issn.2095-1353.2003.004 |
李进, 陈可咏, 李渤生, 等. 1998. 不同海拔高度川滇高山栎群体遗传多样性的变化[J]. 植物学报, 40(8): 761-767. |
李竟, 房守敏, 刘文明, 等. 2005. SSR标记在中国野桑蚕和家蚕的遗传多态性分析中的应用[J]. 蚕业科学, 31(3): 251-256. |
刘宁. 2003. 亚洲玉米螟不同地理种群生态适应性和种群遗传结构的RAPD分析[J]. 中国农业科学院硕士学位论文. |
乔传令, 王靖, 黄菁. 2000. 不同地理种群的小菜蛾的抗性及抗性基因频率[J]. 农药学学报, 2(3): 25-29. |
沈利. 2004. 家蚕SSR标记和CAPS标记的筛选及其应用[J]. 中国科学院硕士学位论文. |
魏建荣, 杨忠岐, 苏智. 2003. 利用生命表评价白蛾周氏啮小蜂对美国白蛾的控制作用[J]. 昆虫学报, 46(3): 318-324. |
薛进, 苏建伟, 黎家文. 2007. 中国水稻二化螟5个地理种群遗传差异的RAPD分析[J]. 湖南农业大学学报:自然科学版, 33(2): 160-163. |
杨忠岐, 张永安. 2007. 重大外来入侵害虫——美国白蛾生物防治技术研究[J]. 昆虫知识, 44(4): 465-471. DOI:10.7679/j.issn.2095-1353.2007.107 |
袁一杨, 高宝嘉, 李明. 2008. 不同林分类型下油松毛虫(Dendrolimus tabulaeformis Tsai et Liu)种群遗传多样性[J]. 生态学报, 28(5): 2099-2106. |
张爱兵, 张真, 王鸿斌. 2004. 中国枯叶蛾科昆虫地理分布及其与环境的相关性[J]. 北京林业大学学报, 26(4): 54-60. |
朱丽虹, 金传玲, 戚凯. 1998. 人工合成美国白蛾性信息素的应用研究[J]. 昆虫知识, 35(4): 225-227. |
Chiharu Koshio, Miwako Tomishima, Koji Shimizu, et al. 2002. Microsatellites in the gypsy moth, Lymantria dispar L.(Lepidoptera: Lymantriidae)[J]. Appl Entomol Zool, 37(2): 309-312. DOI:10.1303/aez.2002.309 |
Ji Yajie, Hua Yueping, Liu Yudi, et al. 2005. Ten polymorphic microsatellite markers developed in the masson pine moth Dendrolimus punctatus[J]. Molecular Ecology Notes, 5: 911-913. DOI:10.1111/men.2005.5.issue-4 |
Lee C E. 2002. Evolutionary genetics of invasive species[J]. Trends in Ecology & Evolution, 8(17): 386-391. |
Meekins J F, Ballard H E, McCarthy B C. 2001. Genetic variation and molecular biogeography of a North American invasive plant species (Alliaria petiolata, Brassicaceae)[J]. International Journal of Plant Science, 162(1): 161-169. DOI:10.1086/317903 |
Mooney H A, Cleland E E. 2001. The evolutionary impact of invasive species[J]. Proceedings of the National Academy Sciences of the United States of America, 98: 5446-5451. DOI:10.1073/pnas.091093398 |
Nissen Scott J, Masters Robert A, Lee Donald J, et al. 1995. DNA-based marker systems to determine genetic diversity of weedy species and their application to biocontrol[J]. Weed Science, 3(43): 504-513. |
Sakai A K, Allendorf F W, Holt J S, et al. 2001. The population biology of invasive species[J]. Annual Review of Ecology and Systematics, 32: 305-332. DOI:10.1146/annurev.ecolsys.32.081501.114037 |
Taylor D R, Aarssen L W. 1990. Complex competitive relationships among genotypes of three perennial grasses: implications for species coexistence[J]. Amr Natur, 136: 105-327. |