 2010, Vol. 46
2010, Vol. 46文章信息
- 杜秀娟, 宋丽文, 高长启, 周春艳, 李兴鹏, 毛宝居, 高群宗, 潘丽铭
- Du Xiujuan, Song Liwen, Gao Changqi, Zhou Chunyan, Li Xingpeng, Mao Baoju, Gao Qunzong, Pan Liming
- 松果梢斑螟不同危害期红松挥发性物质及与寄主选择的关系
- Volatiles from Pinus koraiensis at Different Stages after Infested by Dioryctria pryeri and the Relationship with Host Selection
- 林业科学, 2010, 46(8): 106-113.
- Scientia Silvae Sinicae, 2010, 46(8): 106-113.
-
文章历史
- 收稿日期:2009-04-02
- 修回日期:2009-06-04
-
作者相关文章



2. 吉林省林业科学研究院 长春 130033;
3. 吉林省露水河林业局 抚松 134506
2. Jilin Provincial Academy of Forestry Sciences Changchun 130033;
3. Lushuihe Forestry Bureau of Jilin Province Fusong 134506
松果梢斑螟(Dioryctria pryeri)是我国东北三省红松(Pinus koraiensis)母树林、种子园及人工林中重要害虫,该虫以幼虫蛀食红松球果及枝梢,常造成球果干缩、枝梢枯死,引起严重的经济损失。松果梢斑螟1年1代,其幼虫具有转移危害习性,当原寄生组织营养物质不能满足其生长发育时,便会吐丝随风飘荡或爬行转移危害其他组织。越冬幼虫转移危害期与成虫产卵期,松果梢斑螟危害红松球果; 而越冬幼虫转移危害期与幼虫转移越冬期,其危害红松松塔果痕下面2年生枝(毛宝居等,2006)。植食性昆虫与植物在长期协同进化过程中,形成了特定寄主植物范围,某一虫期或几个虫期取食或产卵于寄主植物的特定组织与器官(尹姣等,2005); 而在寄主选择或识别过程中,寄主植物释放的挥发性物质起着重要通讯引导作用(Kessler et al., 2001; Frédéric et al., 2004)。昆虫通过感受寄主特定的化学指纹图对寄主进行定向与识别(刘勇等,2001; Mumm et al., 2004)。目前,有研究分析了松果梢斑螟寄主红松、油松(P.tabulaeformis)和华山松(P.armandii)的挥发性物质(李新岗等,2006; 2009; 苏晓雨等,2006),并测定了油松和华山松挥发性物质中引诱松果梢斑螟的成分(李新岗等,2006; 2009),未见松果梢斑螟越冬幼虫转移危害期红松不同组织、成虫产卵期红松球果挥发性物质分析及其危害红松机制的研究报道。为此,本文分析了松果梢斑螟越冬幼虫转移危害期红松健康球果、受害2年生枝、健康2年生枝、嫩梢、针叶及成虫产卵期健康与受害红松球果挥发性物质成分与含量,并利用Y型嗅觉仪测试了越冬幼虫对越冬幼虫转移危害期红松不同组织主要挥发性物质标样乙醇溶液的选择,及对红松健康球果主要挥发性物质及其单、双组分标样乙醇溶液选择行为反应,以期为该虫综合治理提供理论基础。
1 材料与方法 1.1 供试昆虫与植物松果梢斑螟幼虫采集于露水河红松种子园或红松母树林。幼虫转移越冬期,大量采摘红松松塔时,采集幼虫饲养于含有红松球果果鳞和果轴碎屑的培养盆中,定期喷水保湿; 当80%幼虫结茧准备越冬时,于4 ℃冰箱中保存4天后,室温下保存2天,即为模拟越冬幼虫,用于越冬幼虫嗅觉反应测试。
吉林省露水河林业局红松种子园内,选择林龄相同、结实量大、生长势良好红松样树10株,于5月中旬(松果梢斑螟越冬幼虫转移危害期),收集红松健康球果、受害2年生枝、健康2年生枝、嫩梢及针叶挥发性物质; 6月下旬(松果梢斑螟成虫产卵期)选取球果未受松果梢斑螟危害样树3株,收集红松健康球果挥发性物质; 7月初(松果梢斑螟初孵幼虫危害期),10株样树中选取球果被松果梢斑螟初孵幼虫危害样树3株,收集红松受害球果挥发性物质。
1.2 植物挥发性物质分析利用大气采样仪收集红松挥发性物质,以0.14 g Tenax-TA吸附剂玻璃管进行动态活体顶空采样。晴朗天气10:00—14:30左右采样15 min,大气采样仪空气流量0.5 L·min-1。取样后样品管放入硅胶干燥器中待用(王鸿斌等,2005; 胡永建等,2007; 任琴等,2005)。
1.3 TCT-GC/MS分析挥发性物质的分离鉴定利用TCT-GC/MS进行分析。仪器工作条件如下,TCT(型号:CP-4010 PTI/TCT)载气压力20 kPa,进样口温度250 ℃(10 min),冷井温度-100 ℃(3 min),脱附温度250 ℃,冷井时进样温度260 ℃; GC(型号:TRACETM 2000)色谱柱DB-5 Low Bleed/MS柱(60 m×0.32 mm× 0.5 μm),液氮脱附进样,He载气,程序升温40 ℃(3 min)后6 ℃·min-1的速率升至270 ℃(5 min),柱后升温至280 ℃(5 min); MS(型号:VOYAGER MASS)EI源,电子能量70 eV,质量范围29~350 u,接口温度250 ℃,离子源温度200 ℃,灯丝电流150 μA,全扫描,扫描速度0.4 s·scan-1。各成分通过与NIST 98图谱库(Xcalibur 1.2)标准化合物的质谱图核对并分析后进行定性,通过面积归一化法对挥发性物质成分以相对含量定量。
1.4 越冬幼虫的嗅觉反应Y-型嗅觉仪测定松果梢斑螟越冬幼虫趋性反应。Y-型嗅觉仪的适应臂长25 cm,两选择臂长均20 cm,选择臂夹角为120°。放置刺激样品锥形瓶陷阱分别与两选择臂相连。此装置两选择臂以大气采样仪为动力,空气分别经活性碳过滤后进入气味源和对照臂陷阱后,进入各自选择臂,汇集于适应臂后排到大气中。为消除不均匀光强对测试结果的影响,在嗅觉仪正上方放置1个10 W照明灯,使两选择臂光强保持一致。管内空气流速为1 L·min-1,连续将6头待测幼虫通过放虫孔放入适应臂,观察并记录其对两选择臂选择情况。幼虫进入锥形瓶陷阱,并停留3 min以上者,记为选择与该臂相连的引诱源; 幼虫不选择两选择臂或进入某一选择臂后又返回,记为无反应。每1样品重复6次,共测试36头幼虫。测试另一样品时,嗅觉仪用大气采样仪连续吹气5 min后,无水乙醇冲洗,然后用电吹风吹干。
气味源为越冬幼虫转移危害期特定组织挥发性物质相对含量标样乙醇溶液; 此时,分析的红松不同组织主要挥发性物质大部分为a-蒎烯、β-蒎烯和柠檬烯,故对不同组织a-蒎烯(98.0%,Aldrich公司)、β-蒎烯(>94.0%,Fisher公司)、柠檬烯(≥96.0%,Fluka公司)各自百分含量混合物、单组分或双组分乙醇溶液进行测试(百分含量详见表 2)。取每种溶液20 μL于定性滤纸上作为气味源。
|
|
采用SPSS13.0分析软件。单因素(ANOVA)F检验(LSD)比较松果梢斑螟不同危害期3种类型红松球果或越冬幼虫转移危害期红松不同组织相同挥发性物质成分相对含量的差异显著性; 成对样本t检验,比较越冬幼虫对两选择臂选择百分率差异。
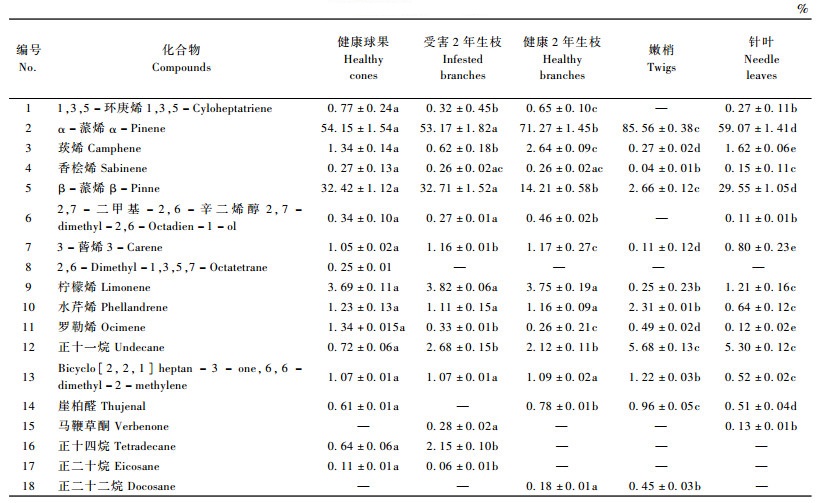
2 结果与分析 2.1 松果梢斑螟寄主红松球果挥发性物质松果梢斑螟越冬幼虫转移危害期,红松健康球果分离鉴定出结构的挥发性物质16种,其中萜类化合物12种、直链烃类3种、环烯烃1种,主要成分为α-蒎烯(54.15%)、β-蒎烯(32.42%)和柠檬烯(3.69%)(表 1); 而其成虫产卵期红松健康和受害球果分离鉴定出结构的挥发性物质均10种,其中萜类化合物6种、直链烃类物质4种,健康球果挥发性物质的主要成分为α-蒎烯(52.62%)、4(10)-崖柏烯(3.30%)和柠檬烯(38.62%),而受害球果挥发性物质的主要成分为α-蒎烯(55.24%)、β-蒎烯(26.74%)和柠檬烯(12.12%)。
|
|
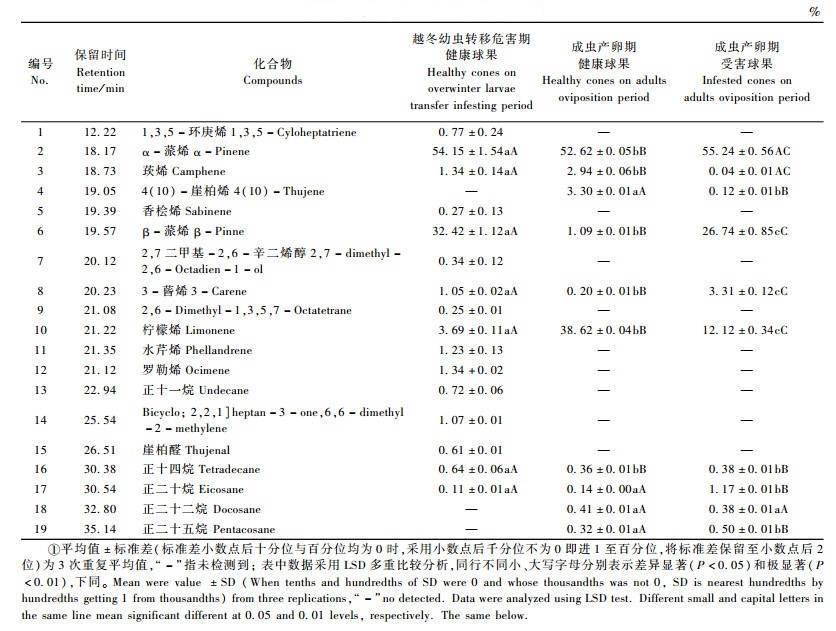
3种类型球果中相同挥发性物质7种,分别为α-蒎烯、柠檬烯、β-蒎烯、3-蒈烯、莰烯、正十四烷和正二十烷,而5种萜类化合物的含量在不同类型球果之间存在着显著或极显著差异,且2种烷烃类物质的含量在越冬幼虫转移危害期健康球果与成虫产卵期球果间差异显著或极显著(表 1)。两时期球果均具有特异性挥发性物质,1, 3, 5-环庚烯、香桧烯、2, 7-二甲基-2, 6-辛二烯醇、2, 6-Dimethyl-1, 3, 5, 7-Octatetrane、水芹烯、罗勒烯、崖柏醛和正十一烷8种挥发性物质仅发现于越冬幼虫转移危害期红松健康球果,而4(10)-崖柏烯、正二十二烷和正二十五烷则仅产生于成虫产卵期红松球果。
2.2 越冬幼虫转移危害期红松不同组织挥发性物质松果梢斑螟越冬幼虫转移危害期红松不同组织挥发性物质分析发现(表 2),越冬幼虫转移危害期,红松不同组织挥发性物质组成不同,其释放的挥发性物质种类由高到低的顺序依次为健康球果、受害2年生枝、健康2年生枝、针叶和嫩梢,分别为16, 15, 14, 14和12种; 而挥发性萜类化合物分别为12, 11, 12, 12和10种,且各自总含量均在90%以上。不同组织产生挥发性物质的主要成分大部分为α-蒎烯、β-蒎烯和柠檬烯。
越冬幼虫转移危害期,红松不同组织的相同挥发性物质包括α-蒎烯、莰烯、香桧烯、β-蒎烯、3-蒈烯、柠檬烯、水芹烯、罗勒烯、正十一烷和Bicyclo [2, 2, 1]heptan-3-one, 6, 6-dimethyl-2-methylene。红松健康球果、健康2年生枝与受害2年生枝释放的相同挥发性物质含量大部分差异不显著,而健康球果、嫩梢及针叶产生的相同挥发性物质含量大都显著不同; 此外,健康2年生枝与受害2年生枝、嫩梢与针叶释放的相同挥发性物质大部分含量差异达显著水平。仅红松嫩梢不产生1, 3, 5-环庚烯。崖柏醛、马鞭草酮、正十四烷、正二十烷和正二十二烷仅在红松某些组织中发现:健康2年生枝、受害2年生枝、嫩梢和针叶释放醛酮类物质,其中健康2年生枝、嫩梢和针叶产生崖柏醛,而受害2年生枝和针叶释放马鞭草酮; 健康球果、健康2年生枝、受害2年生枝和嫩梢释放产生烷烃类物质,其中健康球果和受害2年生枝产生正十四烷、正二十烷,而健康2年生枝和嫩梢产生正二十二烷。
2.3 松果梢斑螟越冬幼虫的嗅觉反应 2.3.1 越冬幼虫对红松不同组织挥发性物质标样乙醇溶液嗅觉反应松果梢斑螟模拟越冬幼虫对越冬幼虫转移危害期红松不同组织挥发性物质标样乙醇溶液的嗅觉反应测试发现(图 1-a),模拟越冬幼虫对红松健康2年生枝、针叶及健康球果主要挥发性物质标样乙醇溶液的选择明显高于乙醇对照组,而对嫩梢及受害2年生枝主要挥发性物质标样乙醇溶液与乙醇对照组的选择差异不显著。

|
图 1 模拟越冬幼虫对红松不同组织主要挥发性物质标样乙醇溶液的嗅觉反应 Figure 1 Olfactory responses of simulated overwinter larvae to alcohol solution with standard sample of main volatile components from different tissues of P. koraiensis *, **: P<0.05,P<0.01; HB, CE, TG, NE和DB分别表示健康2年生枝、健康球果、嫩梢、针叶和受害2年生枝的挥发性物质标样乙醇溶液,AA为无水乙醇,N表示越冬幼虫对气味源无选择。下同。 HB, CE, TG, NE and DB mean alcohol solution with the standard sample of main volatiles from healthy branches, healthy cones, twigs, needles, and damaged branches of P. koraiensis. AA means absolute alcohol, and N means no choice. The same below. |
松果梢斑螟模拟越冬幼虫对红松健康2年生枝、针叶及健康球果挥发性物质标样乙醇溶液的选择测试发现(图 1-b),模拟越冬幼虫对健康球果主要挥发性物质标样乙醇溶液选择的百分率高于针叶及健康2年生枝主要挥发性物质标样乙醇溶液,而且显著高于针叶主要挥发性物质标样乙醇溶液。可见,松果梢斑螟模拟越冬幼虫显著趋向于红松健康球果主要挥发性物质标样乙醇溶液。
2.3.2 越冬幼虫对红松健康球果挥发性物质及其单组分、双组分标样乙醇溶液嗅觉反应松果梢斑螟模拟越冬幼虫对红松健康球果主要挥发性物质标样乙醇溶液及其单组分、双组分标样乙醇溶液的选择行为测试发现(图 2-a),其对α-蒎烯、柠檬烯及α-蒎烯+β-蒎烯乙醇溶液的选择显著或极显著高于健康球果主要挥发性物质标样乙醇溶液,而对β-蒎烯+柠檬烯乙醇溶液的选择显著低于健康球果主要挥发性物质标样乙醇溶液。

|
图 2 模拟越冬幼虫对红松健康球果、单组分及双组分挥发性物质标样乙醇溶液的嗅觉反应 Figure 2 Olfactory responses of simulated overwinter larvae to alcohol solution with standard sample compounds of main volatiles, single and double components from healthy cones of P. koraiensis α, β, L, αβ, αL和βL分别表示红松健康球果主要挥发性物质中α-蒎烯、β-蒎烯、柠檬烯、α-蒎烯+β-蒎烯、α-蒎烯+柠檬烯和β-蒎烯+柠檬烯各百分含量标样的乙醇溶液。 α, β, L, αβ, αL and βL mean alcohol solution with standard sample of α-pinene, β-pinene, limonene, α-pinene+β-pinene, α-pinene+limonene, and β-pinene+limonene, which has percent of volatiles from cones of P. koraiensis, respectively. |
松果梢斑螟模拟越冬幼虫对红松健康球果α-蒎烯、柠檬烯和α-蒎烯+β-蒎烯乙醇溶液的嗅觉反应测试表明(图 2-b),模拟越冬幼虫对α-蒎烯乙醇溶液的选择极显著高于柠檬烯乙醇溶液,而对柠檬烯乙醇溶液的选择显著高于α-蒎烯+β-蒎烯乙醇溶液。说明松果梢斑螟越冬幼虫转移危害期,其选择红松寄主时α-蒎烯起主要作用。
3 结论与讨论相对于松果梢斑螟成虫产卵期红松健康球果而言,本研究中未发现该时期红松受害球果挥发性萜类化合物特异性组分,与CH2Cl2提取法发现松果梢斑螟该时期寄主油松和华山松受害球果具特异性组分不同(李新岗等,2006)。此时,红松受害球果α-蒎烯与β-蒎烯含量高于健康球果,这与D. horneana寄主植物P. cubensis受害树α-蒎烯与β-蒎烯含量高于健康树的结论相似(Irena et al., 1995)。曾有研究发现,α-蒎烯在南方松球果螟(D. amatella)雌蛾产卵位置识别时起重要作用(Fatzinger et al., 1985; Fidgen et al., 1996; Hanula et al., 1985; 李新岗等,2006),而松果梢斑螟成虫产卵期,红松受害球果α-蒎烯含量高于健康球果,可推测松果梢斑螟成虫趋向于红松受害球果产卵。这与松果梢斑螟成虫产卵于油松受害组织的生物学特性一致(李新岗等,2006); 也与昆虫成虫产卵具有“印痕”现象,一般趋向于具有幼虫部位产卵的结论相符。赵成华等(2003)发现β-蒎烯含量高的针叶可降低马尾松毛虫(Dendrolimus punctatus)产卵嗜好,松果梢斑螟成虫产卵期红松受害球果β-蒎烯含量显著高于健康球果,可推测该虫趋向于红松健康球果产卵; 此外,红松健康球果单萜成分含量高于受害球果,依据南方松球果螟明显趋向单萜成分含量高的组织产卵习性(Fatzinger et al., 1985; Fidgen,1996; Hanula,1985; 李新岗,2006),也可推测松果梢斑螟产卵于红松健康球果。α-蒎烯、β-蒎烯与单萜类含量的优势性均决定松果梢斑螟产卵球果类型。而松果梢斑螟成虫较难获得,故一直未进行其成虫产卵时选择红松球果类型的生物测定。
松果梢斑螟越冬幼虫转移危害期,红松球果组织幼嫩,次生代谢活动旺盛,因而红松健康球果挥发性萜类化合物种类较多。越冬前,松果梢斑螟幼虫取食寄生组织时,受害球果低挥发性萜类化合物及果实特性在近距离范围内起作用,而球果营养可能在球果适合度上起作用(Chapman,2003; 李新岗等,2006)。随着气候转冷及红松种子成熟对球果营养物质的大量需求,寄生球果营养不能满足松果梢斑螟幼虫生长需求时,松果梢斑螟转移至球果果痕下2年生枝越冬。松果梢斑螟不同发育阶段及寄主红松不同生理状态下,红松球果的挥发性萜类化合物协调释放,为该虫对红松定向、产卵、逃避和取食等发挥作用(杜家纬,2001; 李新岗等,2006; Mumm et al., 2004; 2005)。
针叶树不同组织产生的挥发性物质组成不同且某些组织具有特异性成分(Kristina et al., 1995),而松果梢斑螟越冬幼虫转移危害期,除红松不同组织挥发性物质组成不同外,某些组织具特异挥发物性物质。模拟越冬幼虫对越冬幼虫转移危害期红松健康2年生枝、针叶及健康球果主要挥发性物质标样乙醇溶液的选择明显高于乙醇对照组,显著趋向于红松健康球果主要挥发性物质标样乙醇溶液,这与其该时期主要危害红松健康2年生枝的生物学特性不同。究其原因,松果梢斑螟幼虫越冬组织——红松2年生枝营养物质不能满足越冬幼虫生长化蛹需求时,转移危害红松其他组织(Chapman,2003),此时红松组织高挥发性萜类化合物发挥重要作用。本研究采用Tenax-TA吸附剂及TCT-GC/MS提取分析红松挥发性物质,测定的主要为红松不同组织自然状态下高挥发性物质,在昆虫远距离寄主定向中发挥作用。此时,红松健康球果挥发性物质可能在松果梢斑螟越冬幼虫远距离定向寄主过程中发挥作用,而健康2年生枝挥发性物质及营养物质则在近距离适合度上发挥作用; 这与松果梢斑螟越冬幼虫转移危害时,大都危害具红松健康球果2年生枝的生物学特性一致。但不排除松果梢斑螟模拟越冬幼虫与越冬幼虫生理上差异对结果的影响。
松果梢斑螟模拟越冬幼虫对红松健康球果的α-蒎烯、柠檬烯、α-蒎烯+β-蒎烯各百分含量乙醇溶液的选择显著高于红松健康球果主要挥发性物质标样乙醇溶液,且这3者对其引诱强弱顺序为α-蒎烯、柠檬烯及α-蒎烯+β-蒎烯乙醇溶液。α-蒎烯乙醇溶液对模拟越冬幼虫的引诱作用最强,这与α-蒎烯在南方松球果螟D. amatella成虫产卵选择寄主时发挥重要作用一致(Fatzinger et al., 1985)。β-蒎烯含量高松树对马尾松毛虫幼虫抗性强(任琴等,2005),β-蒎烯对蛾类幼虫具有趋避作用; 松果梢斑螟模拟越冬幼虫对β-蒎烯或β-蒎烯+柠檬烯乙醇溶液选择百分率低于红松健康球果主要挥发性物质标样乙醇溶液,可推测β-蒎烯对松果梢斑螟模拟越冬幼虫具有趋避作用。α-蒎烯+β-蒎烯乙醇溶液对模拟越冬幼虫的引诱作用显著高于红松健康球果主要挥发性物质标样乙醇溶液; 此时红松健康球果α-蒎烯的含量显著高于β-蒎烯,模拟越冬幼虫转移危害时,α-蒎烯起主导作用,而β-蒎烯仅起辅助调节作用,故β-蒎烯的趋避作用对诱蛾效果影响不大。将来,可于松果梢斑螟越冬幼虫转移危害期,利用该时期红松健康球果α-蒎烯、柠檬烯或α-蒎烯+β-蒎烯含量乙醇溶液干扰松果梢斑螟越冬幼虫对寄主红松的定向,从而控制其对红松的危害。
杜家纬. 2001. 植物-昆虫间的化学通讯及其行为控制[J]. 植物生理学报, 27(3): 193-200. |
胡永建, 任琴, 金幼菊, 等. 2007. 马尾松、湿地松挥发性化学物质的昼夜节律释放[J]. 生态学报, 27(2): 565-570. |
李新岗, 刘惠霞, 刘拉平, 等. 2006. 影响松果梢斑螟寄主选择的植物挥发物成分研究[J]. 林业科学, 42(6): 72-78. |
李新岗, 杨立军, 刘拉平, 等. 2009. 松果梢斑螟成虫的寄主选择[J]. 林业科学, 45(2): 75-81. DOI:10.11707/j.1001-7488.20090213 |
刘勇, 胡萃, 倪汉祥, 等. 2001. 不同营养层次挥发物对燕麦蚜茧蜂寄主搜寻行为的影响[J]. 应用生态学报, 12(4): 581-584. |
毛宝居, 周胜利, 徐清山, 等. 2006. 果梢斑螟生物学特性的研究[J]. 吉林林业科技, 35(2): 29-31. |
任琴, 李镇宇, 胡永建, 等. 2005. 受害马尾松、湿地松挥发性化学物质的释放[J]. 生态学报, 25(11): 2928-2932. DOI:10.3321/j.issn:1000-0933.2005.11.019 |
苏晓雨, 王静, 杨鑫, 等. 2006. 气相色谱-质谱技术分析红松松塔挥发性成分[J]. 分析化学, 34(S1): 217-219. |
王鸿斌, 张真, 孔祥波, 等. 2005. 油松萜烯类挥发物释放规律与红脂大小蠹危害的关系[J]. 北京林业大学学报, 27(2): 75-80. |
尹姣, 曹雅忠, 罗礼智, 等. 2005. 草地螟对寄主植物的选择性及其化学生态机制[J]. 生态学报, 25(8): 1844-1852. |
赵成华, 阎云花. 2003. 马尾松针叶中的挥发物质对马尾松毛虫产卵行为的影响[J]. 林业科学, 39(6): 91-93. DOI:10.11707/j.1001-7488.20030616 |
Chapman R F. 2003. Contact chemoreception in feeding by phytophagous insects[J]. Annual Review of Entomology, 48: 455-484. DOI:10.1146/annurev.ento.48.091801.112629 |
Fatzinger C W, Merkel E P. 1985. Oviposition and feeding preferences of the southern pine coneworm for different host-plant-materials and observations on monoterpenes as an oviposition stimulant[J]. Journal of Chemical Ecology, 11(6): 689-699. DOI:10.1007/BF00988298 |
Frédéric F, Georges L, Eric H. 2004. Olfatory responses to aphid and host plant volatile releases: (E)-β-Farnesene an effective kairomone for the predator Adalia bipmctata[J]. Journal of Chemical Ecology, 30(4): 741-755. DOI:10.1023/B:JOEC.0000028429.13413.a2 |
Fidgen L, Sweeney J. 1996. Fir coneworms, Diorytria abietivorella, prefer cons reviously exploited by spruce cone maggots, Strobilomyia neatithracina Michelsen and S. appalachensis Michelsen. IUFRO Working Party: S7. 03-01 Cone and Seed Insects, 5-6.
|
Hanula J L, Berisford C W, Debarr G L. 1985. Monoterpene oviposition stimulants of Diorytria amatella in volatiles from fusiform rust galls and second-year loblolly pine cones[J]. Journal of Chemical Ecology, 11(7): 943-952. DOI:10.1007/BF01012080 |
Irena V, Kristina S, Jan V, et al. 1995. Contents and enantiomeric composition of monoterpene hydrocarbons in xylem oleoresins from four Pinus species growing in Cuba[J]. Comparison of trees unattacked and attacked by Dioryctria horneana. Biochemical Systematic and Ecology, 23(1): 1-15. |
Kessler A, Baldwin I T. 2001. Defensive function of herbivore-induced plant volatile emissions in nature[J]. Science, 291: 2141-2144. DOI:10.1126/science.291.5511.2141 |
Kristina Sjödin, Monika Persson, Anna-Karin Börg-Karlson, et al. 1995. Enatiomeric compositions of monoterpene hydrocarbons in different tissues of four individuals of Pinus sylvestris[J]. Phytochemistry, 41(2): 439-445. |
Mumm R, Hilker M. 2005. The significance of background odour for an egg parasitoid to detect plants with host eggs[J]. Chemical Senses, 30(4): 337-343. DOI:10.1093/chemse/bji028 |
Mumm R, Tiemann T, Schulz S, et al. 2004. Analysis of volatiles from black pine (Pinus nigra): significance of wounding and egg deposition by a herbivorous sawfly[J]. Phytochemistry, 659(24): 3221-3230. |