 2010, Vol. 46
2010, Vol. 46文章信息
- 吴若菁, 庄捷, 陈清松, 陈文萍
- Wu Ruojing, Zhuang Jie, Chen Qingsong, Chen Wenping
- 铅胁迫下马尾松微核率与生理指标变化的相关分析
- Correlations between of Micronucleus Rate and Physiological Changes of Pinus massoniana in Response to Lead Stress
- 林业科学, 2010, 46(8): 78-83.
- Scientia Silvae Sinicae, 2010, 46(8): 78-83.
-
文章历史
- 收稿日期:2009-10-19
- 修回日期:2010-06-02
-
作者相关文章


2. 福建商贸学校 福州 350007;
3. 福建师范大学化学与材料学院 福州 350007
2. Fujian Business School Fuzhou 350007;
3. College of Chemistry and Materials Sciences, Fujian Normal University Fuzhou 350007
铅是五毒元素之一,多与锌矿、铜锌矿伴生。随着矿产资源的开发,铅对山地土壤的污染和对植物的毒害作用日益严重,有关铅胁迫对草本植物和农作物生理生化特性的影响以及抗性机制的研究,国内外已有不少报道(Kupper et al., 1996; Morelli et al., 2001; 詹洁等,2009; 姚广等,2009; 谢传俊等,2008),而有关林木方面则研究较少(王连芳等,2009)。研究重金属对林木的影响和筛选抗性较强的树种,通过造林达到既绿化荒山又能吸收、富集山地土壤中的重金属,修复受污染山地的目的。马尾松(Pinus massoniana)是我国南方荒山的先锋造林树种和主要用材林树种,但是马尾松对铅胁迫的遗传变异和生理生化变化,以及这些指标间的变化相关性,目前还尚未见到报道。本文拟开展这方面的研究,为马尾松在污染矿区的荒山造林和进一步开展马尾松的抗性选择育种提供相关的试验依据。
1 材料与方法 1.1 马尾松苗木的培育马尾松种子由福建省林业厅种苗站提供。醋酸铅采用国产分析纯。选择饱满、大小均匀的马尾松种子浸种24 h,播种于湿沙中(沙子先用3%HCl溶液浸泡1星期,用流水冲洗干净,再用去离子水漂洗后烘干)24 ℃恒温催芽,中间酌情补充水分。将萌发的种子和幼苗按照试验的不同要求分别进行胁迫处理,幼苗处理期间每隔3天浇1次Hoagland营养液(连兆煌,2002)。所有试验的每个处理重复3次,对照均采用去离子水处理。
1.2 试验方法 1.2.1 铅胁迫下马尾松根尖微核率的观察用去离子水配制浓度为100,1 000,2 000,3 000 mg·L-1的单一醋酸铅溶液。待马尾松种子幼根长至约0.5~0.8 cm时,进行不同的胁迫处理24 h,微核率的测定采用吴若菁等(2007)的方法。每个处理随机观察2 000个左右的细胞,3次重复。采用YS100型Nikon显微镜进行观察。
1.2.2 铅胁迫下马尾松幼苗生理生化指标变化的测定待苗高4 cm时挖取长势一致的马尾松幼苗移植于统一规格的花盆中,用去离子水将醋酸铅配制成不同浓度处理液,使沙培盆栽中铅的污染水平分别处于: 100,1 000,2 000,3 000 mg·L-1,对马尾松幼苗进行胁迫处理。每盆每隔24 h喷洒1次处理液,每隔3天浇1次Hoagland营养液,处理7天后,取样测定各项生理生化指标,每处理测定3次。对照采用去离子水培养幼苗。
MDA含量的测定采用硫代巴比妥酸(TBA)显色法(王晶英等,2003); 质膜透性的测定采用电导率仪法,伤害度=(处理电导率-空白电导率)/(煮沸电导率-空白电导率)(李合生等,2001); SOD活性的测定采用氮蓝四唑(NBT)还原法(李合生等,2001); POD活性的测定采用愈创木酚法(张志良等。2003)。
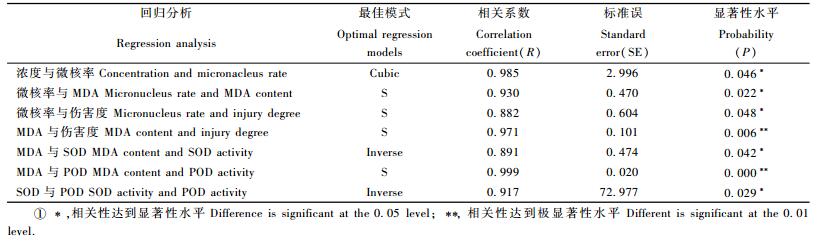
1.2.3 数据处理与分析试验数据采用SPSS13.0软件进行方差分析和t检验,并进行浓度、微核率、MDA、伤害度、POD和SOD之间的相互回归分析。采用的回归模式为:线性方程(Linear),二次曲线(Quadratic),混合曲线(Compound),生长曲线(Growth),三次曲线(Cubic),S曲线(S),指数曲线(Exponential),反曲线(Inverse),幂函数(Power),拟合Logistic曲线模块(Logistic),对数曲线(Logarithmic)。最后根据实际观测值与不同模式回归的吻合结果,筛选出最佳的回归方程。
2 结果与分析 2.1 胁迫条件下马尾松幼苗根尖细胞的微核率变化在遗传毒理学研究中,细胞微核率常用作评价各种物质对染色体损伤的指标(李龙等,2006)。试验结果表明:在铅胁迫下,马尾松幼苗根尖细胞有的会产生染色体端部断裂,进而形成后期桥和断片(图 1,箭头所指为断片)。有的则产生落后染色体(图 2),这些染色体断片和落后染色体最终形成微核(图 3)。随着铅浓度的上升,细胞的微核率急剧上升(图 4),在铅浓度达到3 000 mg·L-1时,微核率达到最高值。对不同浓度的胁迫处理结果进行方差分析,表明铅对微核率的影响(F=135.948,P=0.000)达到极显著差异。此外,将不同处理的结果与对照进行t检验,结果均达到极显著差异。

|
图 1 染色体桥断裂及断片(×600) Figure 1 Chromosome bridge breakage and fragment(×600) |

|
图 2 落后染色体(×600) Figure 2 Lagging chromosomes(×600) |

|
图 3 微核(×600) Figure 3 Micronucleus(×600) |

|
图 4 铅对细胞微核率的影响 Figure 4 Influence of lead on cell micronucleus rate |
MDA含量可反映膜质过氧化作用的强弱,可表示膜受伤害或变性程度(李合生等,2001)。在铅的胁迫下,马尾松幼苗的MDA含量变化如图 5,质膜伤害度如图 6所示。随着铅浓度的上升,马尾松幼苗体内MDA含量不断增多,质膜伤害度也逐渐加大,在2 000 mg·L-1时,2者达到最大值。而当铅浓度>2 000 mg·L-1时,随着铅浓度的增加,MDA含量和伤害度出现不升反降的现象,其原因可能是当铅胁迫对细胞质膜的伤害达到一定程度时,导致质膜的选择性通透屏障被严重破坏,膜体系崩溃,反而出现下降趋势。

|
图 5 铅对马尾松MDA含量的影响 Figure 5 Influence of lead on P. massoniana MDA content |

|
图 6 铅对马尾松细胞伤害度的影响 Figure 6 Influence of lead on P. massoniana damage degree |
对铅胁迫状态下的马尾松幼苗进行SOD和POD测定,结果见图 7,8。可以看出随着铅浓度的升高,马尾松幼苗的SOD和POD活性呈现先升后降,在铅浓度为2 000 mg·L-1时,2者达到最高值,而后随着铅浓度的继续升高,开始下降。其原因可能是铅的胁迫毒害作用超过马尾松的耐受力时,会抑制SOD和POD活性, 所以出现先升后降现象。

|
图 7 铅对马尾松SOD活性的影响 Figure 7 Influence of lead on P. massoniana SOD activity |

|
图 8 铅对马尾松POD活性的影响 Figure 8 Influence of lead on P. massoniana POD activity |
通过采用12种不同的回归模式对各项指标间进行分析,筛选出最佳的回归模式见表 1,图 9~15。
|
|

|
图 9 铅浓度与微核率的Cubic回归模拟 Figure 9 The simulated Cubic regression betweenlead concentration and micronucleus rate |

|
图 10 微核率与MDA间的S回归模拟 Figure 10 The simulated S regression between micronucleus rate and MDA |

|
图 11 微核率与伤害度间的S回归模拟 Figure 11 The simulated S regression between micronucleus rate and cell degree of injury |

|
图 12 MDA与伤害度间的S回归模拟 Figure 12 The simulated S regression between MDA and cell degree of injury |

|
图 13 MDA与SOD间的Inverse回归模拟 Figure 13 The simulated Inverse regression between MDA and SOD |

|
图 14 MDA与POD间的S回归模拟 Figure 14 The simulated S regression between MDA and POD |

|
图 15 SOD与POD间的Inverse回归模拟 Figure 15 The simulated Inverse regression between SOD and POD |
对不同铅胁迫浓度对微核率的影响进行不同模式的回归分析结果发现:当采用Cubic模式时效果最佳(图 9),即Y=4.429+0.006X+2.394×10-6X2-5.6×10-10X3时(Y为微核率,X为浓度),浓度与核异常率之间的相关性r=0.985时,P=0.046,达到显著相关。
2.3.2 铅胁迫下微核率与MDA、伤害度间的回归分析试验分析结果表明:微核率与MDA、伤害度间最佳回归模式均是S模式,当微核率与MDA间采用Y=e4.677-6.466/X(Y为微核率,X为MDA)时,2者的相关性r=0.930,P=0.022,达到显著相关,其模拟回归见图 10。微核率与伤害度采用Y=e9.972-0.928/X时,2者的相关性r=0.882,P=0.048,达到显著相关,其模拟回归见图 11。
2.3.3 铅胁迫下MDA与伤害度、SOD和POD间的回归分析将MDA与伤害度、SOD和POD分别进行不同模式回归分析的结果,它们最佳的回归模式分别是: MDA与伤害度采用S模式,即当Y=e3.853-0.337/X(Y为MDA,X为伤害度)时,它们间的相关性r=0.971,P=0.006 < P=0.01,2者达到极显著相关,其模拟回归见图 12。MDA与SOD采用Inverse模式,即当Y=4.729-58.417/X(Y为MDA,X为SOD)时,它们间的相关性r=0.891,P=0.042 < P=0.05,2者达到显著相关,其模拟回归见图 13。MDA与POD采用S模式,即当Y=e1.964-432.17/X(Y为MDA,X为POD)时,它们间的相关性r=0.999,P=0.000 < P=0.01,2者达到极显著相关,其模拟回归见图 14。
2.3.4 铅胁迫下SOD和POD间的回归分析从不同模式对SOD与POD的回归结果可见2者间的最佳回归模式为Inverse模式,即当Y=834.435-10 491.8/X(Y为SOD,X为POD)时,它们的相关性r=0.917,P=0.029 < P=0.05,2者相关性达到显著,其模拟回归见图 15。
3 讨论 3.1 铅对马尾松根尖细胞染色体的损伤机制本试验的结果表明:铅会使马尾松细胞的遗传物质-染色体产生结构损伤,出现微核。这与前人的报道是相符的(辛晓芸等,2004)。其损伤机制之一是:高浓度的Pb2+能完全竞争出核糖核酸酶中的Ca2+,从而与核糖核酸酶结合,使核糖核酸酶的二级结构遭到严重破坏(洪法水等,2003),降低DNase的活性,并与核酸的碱基结合引起核酸立体结构的变化,使正在分裂的细胞发生染色体畸变(Shahin et al., 1991)。损伤机制之二是: Pb2+的电荷可以诱导植物体内产生大量的·O2,H2O2,·OH等活性氧自由基(Guo et al., 2004),这些活性自由基会对糖残基进行攻击,引起DNA的碎裂、碱基丢失和有末端糖残基片段的链断裂,导致产生上述的染色体结构损伤现象。在本试验中随着铅胁迫的浓度加大微核率逐渐加大,即染色体结构损伤加重的现象。回归分析的结果也进一步证实铅浓度与微核率间的相关性具有显著性。
3.2 铅对马尾松幼苗针叶质膜体系的损伤机制试验结果证明铅的胁迫除了会导致马尾松染色体结构损伤外,还会使马尾松细胞产生质膜损伤,表现出随着铅胁迫浓度增加,MDA含量上升,细胞电导率加大,质膜透性加强。说明在铅胁迫下,产生的大量活性氧会使细胞原生质膜中的不饱和脂肪酸发生过氧化作用产生MDA,MDA伤害脂膜系统,使其选择性被破坏,表现出细胞内电解质外渗量增加,电导率加大,质膜伤害度也随着铅浓度的增加而逐渐加大。但是,当浓度增加到质膜体系被严重破坏时,质膜的选择性通透屏障则失去作用,此时会随着铅浓度增加,MDA和电导率反而下降的现象。此外,活性氧水平升高还会使蛋白质和生物大分子变性,这也是膜损伤的重要原因之一。
对MDA与细胞伤害度间,以及微核率与MDA、细胞伤害度回归分析的结果表明: MDA与细胞伤害度,2者相关性极显著,证明MDA与细胞质膜系统伤害有密切关系。微核率与MDA、细胞伤害度之间的相关性显著,表明染色体损伤导致细胞正常生命活动所需的物质转录受到抑制,影响到了细胞修复功能。因此,当醋酸铅浓度为3 000 mg·L-1时,微核率为最高值,而此时MDA与细胞伤害度却出现下降。
3.3 马尾松幼苗对铅胁迫的抗性机制对马尾松生理生化不同指标的测定结果表明:面对铅胁迫导致的各种损伤,马尾松自身会产生不同的应激反应,启动不同抗性机制,以便进行自我保护。在马尾松的耐受极限范围内,随着胁迫的加重,马尾松体内抗氧化酶系统的机制被激发,超氧化物歧化酶(SOD)活性和过氧化物酶(POD)含量上升。SOD是生物体内防御氧化损伤重要的金属酶之一,其功能是将O2·歧化为H2O2,从而阻止或减少O2·自由基对植株生长的破坏。SOD活性在一定范围内与细胞内的活性氧自由基水平呈正相关(尹永强等,2007)。而POD则利用H2O2来催化对植物自身有毒害过氧化物(POD底物)的氧化和分解以维持自身的正常代谢。对SOD与POD进行回归分析的结果也证明2者间具有显著的相关性。
试验结果还表明:当活性氧水平超过马尾松自身抵御极限时,过多的活性氧会导致严重的遗传物质损伤和质膜系统损伤进而影响到相关酶蛋白的合成。从试验数据中可以看出: SOD和POD活性与MDA和细胞伤害度一样均是在醋酸铅浓度为2 000 mg·L-1时,达到最高值。在浓度为3 000 mg·L-1,微核率处于最高值30.4%,即遗传损伤最大时,出现下降趋势。说明以SOD和POD为代表的抗氧化酶系统对Pb2+毒害的保护作用是有限度的。将MDA与SOD, POD进行回归分析的结果也进一步证实MDA与SOD, MDA与POD间具有显著的相关性。
4 结论1) 单一铅胁迫处理对马尾松微核率的影响,经t检验与对照相比均达到极显著差异。随着铅胁迫浓度的增加,微核率呈现逐渐上升趋势。
2) 在铅胁迫下,随铅浓度的加大,MDA、细胞伤害度、SOD和POD值逐渐上升,在铅浓度为2 000 mg·L-1时达到最高值,超过这个阈值,随着铅浓度的逐渐加大,MDA、细胞伤害度、SOD和POD值开始下降,说明马尾松的自身的抗氧化酶系统对Pb2+毒害的保护作用是有限的。
3) 不同模式的回归分析结果表明:铅浓度与微核率采用Cubic模式; 微核率与伤害度,微核率与MDA,MDA与伤害度,MDA与POD间采用S模式; MDA与SOD,SOD与POD间采用Inverse模式回归时,预测相互间变化效果最佳,相关性可以达到显著或极显著水平。
洪法水, 王玲, 吴康, 等. 2003. Pb2+对核糖核酸酶活性及其结构的影响[J]. 化学学报, 61(1): 117-121. |
李合生. 2001. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社: 167-169, 260-263.
|
李龙, 陈家堏. 2006. 现代毒理学实验技术原理与方法[M]. 北京: 化学工业出版社: 31-35.
|
连兆煌. 2002. 无土栽培原理与技术[M]. 北京: 中国农业出版社: 57.
|
王晶英, 敖红, 张杰. 2003. 植物生理生化实验技术与原理[M]. 哈尔滨: 东北林业大学出版社: 132-138.
|
王连芳, 徐学华, 李玉灵, 等. 2009. 铅胁迫对六种绿化树种叶片生理生化特性的影响[J]. 河北农业大学学报, 32(2): 29-33. |
吴若菁, 陈奋飞, 庄捷, 等. 2007. SO2的衍生物对泥鳅的急性毒性和染色体损伤研究[J]. 水生生物学报, 31(4): 585-589. |
姚广, 高辉远, 王未未, 等. 2009. 铅胁迫对玉米幼苗叶片光系统功能及光合作用的影响[J]. 生态学报, 29(3): 1162-1169. |
尹永强, 胡建斌, 邓明军. 2007. 植物叶片抗氧化系统及其对逆境胁迫的响应研究进展[J]. 中国农学通报, 23(1): 105-110. |
谢传俊, 杨集辉, 周守标, 等. 2008. 铅递进胁迫对假俭草和结缕草生理特性的影响[J]. 草业学报, 17(4): 65-70. |
辛晓芸, 桑楠, 郑国柱. 2004. 铅诱发蚕豆根尖细胞的遗传损伤[J]. 环境与健康杂志, 21(6): 400-401. |
张志良, 瞿伟. 2003. 植物生理学实验指导[M]. 北京: 高等教育出版社: 123-124.
|
Guo T R, Zhang G P, Zhou M X, et al. 2004. Effects of aluminum and cadmium toxicity on growth and antioxidant enzyme activities of two barley genotypes with different Al tolerance[J]. Plant and Soil, 258(1): 241-248. DOI:10.1023/B:PLSO.0000016554.87519.d6 |
Kupper H, Kupper F, Spiller M. 1996. Environmental relevance of heavy metal-substituted chlorophylls using the example of water plants[J]. Exp Bot, 47: 259-266. DOI:10.1093/jxb/47.2.259 |
Morelli E, Scarano G. 2001. Synthesis and stability of numberical chromosomal by cadmium and lead in the marine diatom Phaeodactylum tricornutum[J]. Mar Environ Res, 52: 383-395. DOI:10.1016/S0141-1136(01)00093-9 |
Shahin S A, EI-Amoodi K H. 1991. Induction of numberical chromosomal aberrations during DNA synthesis using the fungicides nimrod and rubigan-4 in root tips of Vicia fata L[J]. Mutat Res, 261: 169-176. DOI:10.1016/0165-1218(91)90064-S |