 2010, Vol. 46
2010, Vol. 46文章信息
- 孙晶, 王庆成, 刘强, 徐静
- Sun Jing, Wang Qingcheng, Liu Qiang, Xu Jing
- NaHCO3胁迫下朝鲜接骨木和茶条槭苗木的生长及生理响应
- Growth and Physiological Responses of Sambucus coreana and Acer ginnala Seedlings to NaHCO3 Stress
- 林业科学, 2010, 46(8): 71-77.
- Scientia Silvae Sinicae, 2010, 46(8): 71-77.
-
文章历史
- 收稿日期:2009-07-10
- 修回日期:2010-06-05
-
作者相关文章



我国盐碱地面积约占国土面积的1/3,由于各种原因导致的土壤次生盐渍化仍在继续(牛冬玲等,2002)。耐盐植物的引选为盐碱地区的环境建设提供重要物质基础。植物耐盐机制和耐盐植物品种的培育已成为当前的研究热点之一(赵可夫,1993)。
现有植物抗盐生理研究多以NaCl为研究对象,少有对碳酸盐胁迫的研究,尤其对林木的碳酸盐抗性的研究更少。植物对盐胁迫的反应是整体的植株反应,必须采用一系列的综合指标来反映植物的抗性(李国雷,2004)。因此,把植物生物膜、酶系统、渗透调节物质、非酶促防御系统、形质指标与生理生化指标等联系起来系统研究树种的耐盐反应特性,进而探讨不同树种不同的生长阶段的抗盐机制,是非常重要的。
朝鲜接骨木(Sambucus coreana)和茶条槭(Acer ginnala)是东北东部山区的乡土树种,树姿优美,季相明显,观赏价值高,在东北地区的城市绿化中得到广泛的应用。目前对朝鲜接骨木和茶条槭多是苗木繁殖和栽培技术方面的研究(张弼弘等,2003; 陈有民,1990),对其耐盐性的研究尚未见报道。本文以朝鲜接骨木和茶条槭1年生苗木为对象,采用温室盆栽的方法,研究不同碳酸盐胁迫强度对苗木生长和生理指标的影响,据此对其耐盐性进行评价。同时,通过对其抗性的研究,确定在盐碱土地区的适生范围,为丰富盐碱地区绿化树种资源奠定理论基础。
1 材料与方法 1.1 试验材料与处理试验于2007年5—9月在东北林业大学帽儿山实验林场尖砬沟森林培育实验站的温室内进行。试验材料为大庆红旗林场苗圃培育的1年生朝鲜接骨木和茶条槭的实生苗,选择大小相对一致、生长正常的苗木用于试验; 栽培容器为底部带有托盘的深色不透明塑料容器(高25 cm,上、下直径分别为30和26 cm); 栽培基质为细河砂,每桶7.5 kg(风干质量)。每盆均匀植苗3株。定植后,每天浇水300 mL,保证基质处于湿润状态。每3天浇1次营养液[按照Utriainen等(2001)方法配制],每盆每次200 mL。定植3个月后苗木缓苗,叶片完全展开时进行盐胁迫处理。
以黑龙江省大庆地区轻、中和重度盐碱土盐含量和pH作为参照(潘保原等,2006),设置盐(NaHCO3)4个水平的浓度处理,即砂土的含盐量分别是0(对照,CK), 1.0 g·kg-1(低盐处理,L),2.0 g·kg-1(中盐处理,I),4.0 g·kg-1(高盐处理,H)。NaHCO3配成7.5 g·L-1溶液,试验开始时各个处理均加入1 L配制好的NaHCO3溶液,至基质中含盐量达到1 g·kg-1为止,以后的含盐量每12 h递增1 g·kg-1,直至达到最高预定含盐量为止,对照浇自来水,每种处理20桶。胁迫期间每天浇适量的自来水以补充散失的水分,营养液正常供给,在盐处理后每隔一段时间将渗到托盘中的盐溶液转回到塑料容器中。于胁迫后第10,20,30天采集苗木中上部叶片混匀置于低温保温箱中带回,用于生理指标的测定。
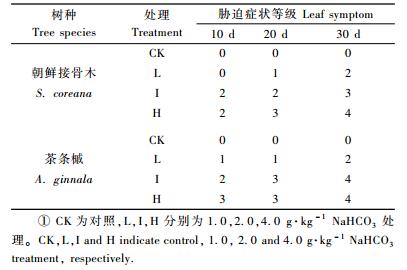
1.2 试验方法 1.2.1 盐胁迫危害症状调查分别于盐胁迫后第10,20,30天观察和记录苗木叶形态以及颜色等的变化,根据危害程度轻重分为5个级别(李国雷,2004),即: 0级:无盐胁迫危害症状; 1级:有少部分叶尖、叶缘和叶脉变黄; 2级:约有1/2的叶尖、叶缘焦枯; 3级:大部分叶片有叶尖、叶缘焦枯和落叶现象; 4级:枝枯、叶落直至死亡。
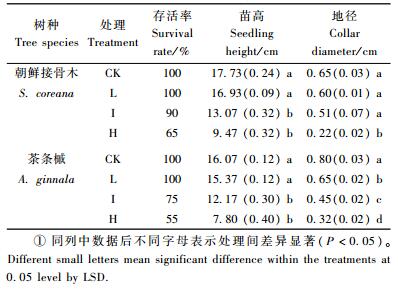
1.2.2 苗木生长指标调查苗高和地径分别在胁迫前、后用钢卷尺和游标卡尺各测定1次,生长量为胁迫前、后2次测量值之差,分别精确到0.1和0.01 cm。胁迫结束时,统计记录苗木的存活株数和死亡株数。苗木存活率=存活苗数/苗木总数×100%。
1.2.3 叶片生理指标测定丙二醛(malondehyde,MDA)采用硫代巴比妥酸(2-thiobarbituric acid,TBA)法测定(李合生等,2000); 相对电导率参照陈建勋等(2002)的方法测定; 超氧化物歧化酶(superoxide dismutase,SOD)活性采用氯化硝基四氮唑蓝光学还原法进行测定(陈建勋等,2002); 过氧化氢酶(catalase,CAT)活性采用紫外吸收法测定(陈建勋等,2002); 谷胱甘肽(glutathione,GSH)和抗坏血酸(ascorbic acid,ASA)含量参照陈建勋等(2002)的方法测定; 脯氨酸含量采用酸性茚三酮比色法(李合生等,2000); 可溶性糖含量采用蒽酮乙酸乙酯法(李合生等,2000)。
1.3 数据分析采用SPSS 13.0(SPSS公司,2005)软件分析进行处理间差异显著性检验,Microsoft Excel(微软公司,2003)制图。
2 结果与分析 2.1 NaHCO3盐胁迫对2树种苗木生长的影响随盐处理浓度的增加,2树种苗木均表现出一定的胁迫症状(表 1),盐胁迫的时间越长,苗木盐害越严重。朝鲜接骨木在低盐处理下,胁迫10天时无明显盐害症状; 胁迫20天后苗木出现明显盐害症状,部分叶尖、叶缘和叶脉变黄; 胁迫30天时盐害症状加剧,约有1/2的叶尖、叶缘焦枯。在中盐处理下,胁迫10和20天时,约有1/2的叶尖、叶缘焦枯,盐害症状明显; 胁迫30天后盐害症状加剧,苗木出现大部分叶片的叶尖、叶缘焦枯和落叶现象。高盐处理下,在胁迫10天时,苗木约有1/2的叶尖、叶缘焦枯,盐害症状明显; 胁迫20天后盐害症状加剧,苗木出现大部分叶片的叶尖、叶缘焦枯和落叶现象; 胁迫30天时,部分苗木死亡。茶条槭在低盐处理下,胁迫10和20天后盐害症状明显,苗木的部分叶尖、叶缘和叶脉变黄; 胁迫30天时盐害症状加剧,约有1/2的叶尖、叶缘焦枯。在中盐处理下,胁迫10天时约有1/2的叶尖、叶缘焦枯,盐害症状明显; 胁迫20天后盐害症状加剧,苗木出现大部分叶片的叶尖、叶缘焦枯和落叶现象; 胁迫30天后,部分苗木枝枯、叶落。高盐处理下,在胁迫10和20天时,苗木出现大部分叶片的叶尖、叶缘焦枯和落叶现象; 30天后植株大部分枝枯、叶落, 部分死亡。
|
|
随盐处理浓度的增加,2树种苗木存活率下降程度不同,苗高、地径的相对生长量均呈显著的下降趋势(表 2)。低盐处理时,2树种成活率未受影响,中、高盐处理时2树种成活率均明显下降,茶条槭下降幅度高于朝鲜接骨木。低盐处理下,朝鲜接骨木的苗高生长量与对照相比无显著差异(P>0.05),中、高盐处理下显著降低(P < 0.05);低、中盐处理下地径生长量与对照无显著差异(P>0.05),高盐处理下显著降低(P < 0.05)。茶条槭的苗高生长量在低盐处理下与对照无显著差异(P>0.05),中、高盐处理下显著降低(P < 0.05),地径相对生长量在各处理上均显著降低(P < 0.05)。
|
|
随盐处理浓度的增大和胁迫时间的延长,2树种苗木叶片MDA含量显著升高(图 1)。胁迫10和20天时,低盐处理与对照间差异不显著(P>0.05),中、高盐处理与对照差异显著(P < 0.05);胁迫30天时,各处理下MDA含量与对照间均差异显著(P < 0.05)。茶条槭苗木叶片的MDA含量在胁迫10天时,各处理与对照间均差异显著(P < 0.05);胁迫20和30天时,低盐处理与对照差异不显著(P>0.05),中、高盐处理与对照间均差异显著(P < 0.05)。

|
图 1 不同时间、不同基质NaHCO3浓度胁迫处理下朝鲜接骨木和茶条槭苗木叶片的MDA含量 Figure 1 MDA contents in S. coreana and A. ginnala seedlings leaves under different concentrations of NaHCO3 salt stress for different days |
随盐处理浓度的增大和胁迫时间的延长,2树种苗木叶片的相对电导率均明显升高(图 2)。朝鲜接骨木在胁迫10天时,低盐处理与对照相比差异不显著(P>0.05),中、高盐处理显著高于对照(P < 0.05),胁迫20天时,各处理均显著高于对照(P < 0.05),胁迫30天时,低、中盐处理与对照相比差异不显著(P>0.05),高盐处理显著高于对照(P < 0.05)。茶条槭在胁迫10和20天时,低、中盐处理与对照相比差异不显著(P>0.05),高盐处理与对照间差异显著(P < 0.05)。胁迫30天时,各处理与对照间均达到显著差异水平(P < 0.05)。

|
图 2 不同时间、不同基质NaHCO3浓度胁迫处理下朝鲜接骨木和茶条槭苗木叶片的相对电导率 Figure 2 Relative electric conductance in S. coreana and A. ginnala seedlings leaves under different concentrations of NaHCO3 salt stress for different days |
胁迫10和20天时朝鲜接骨木叶片的SOD活性,除在20天时的中、高盐处理高于对照外,其余均低于对照,但差异均不显著(P>0.05),胁迫30天时各处理显著低于对照(P < 0.05)。胁迫前20天的茶条槭叶片SOD活性除在10天时低盐处理低于对照外,其他各处理均高于对照,但差异均不显著(P>0.05),胁迫30天时,低盐处理高于对照但未达到显著差异(P>0.05),中、高盐处理则显著低于对照(P < 0.05)(图 3)。盐胁迫下朝鲜接骨木CAT活性在各时期均高于对照,但仅低盐处理在20和30天时达到显著水平(P < 0.05);茶条槭CAT活性在胁迫前20天时除在20天时中盐处理低于对照外,其余均高于对照,但均未达到显著水平(P>0.05),30天时各处理低于对照,中盐处理达显著水平(P < 0.05)(图 4)。

|
图 3 不同时间、不同基质NaHCO3浓度胁迫处理下朝鲜接骨木和茶条槭苗木叶片的SOD活性 Figure 3 SOD activity in S. coreana and A. ginnala seedlings leaves under different concentrations of NaHCO3 salt stress for different days |

|
图 4 不同时间、不同基质NaHCO3浓度胁迫处理下朝鲜接骨木和茶条槭苗木叶片的CAT活性 Figure 4 CAT activity in S. coreana and A. ginnala seedlings leaves under different concentrations of NaHCO3 salt stress for different days |
朝鲜接骨木在各胁迫时期ASA含量随处理浓度的增加整体呈上升趋势,各处理均高于对照,其中在胁迫10天时,高盐处理达显著水平(P < 0.05),20天时各处理均达显著水平(P < 0.05),30天时中盐处理达显著水平(P < 0.05)。随着胁迫时间的延长低盐处理呈先降低后升高的趋势,中、高盐处理呈升高趋势。随盐处理浓度的增加,茶条槭的ASA含量在胁迫10天时,各处理均高于对照,其中高盐处理达显著水平(P < 0.05),胁迫20和30天时,低、中盐处理低于对照,而高盐处理则高于对照,其中低盐处理均达到显著水平(P < 0.05),中、高盐处理与对照间的差异不显著(P>0.05)(图 5)。

|
图 5 不同时间、不同基质NaHCO3浓度胁迫处理下朝鲜接骨木和茶条槭苗木叶片的ASA含量 Figure 5 ASA content in S. coreana and A. ginnala seedlings leaves under different concentrations of NaHCO3 salt stress for different days |
朝鲜接骨木在胁迫10天时各处理的GSH含量均低于对照,其中低、高盐处理达显著水平(P < 0.05);胁迫20和30天时,各处理均高于对照,其中20天时的各处理和30天时的高盐处理达显著水平(P < 0.05)。朝鲜接骨木在胁迫20天时各处理的GSH含量较对照分别增加68.5%, 54.5%, 90.4%。各处理茶条槭在10天时高于对照,其中中盐处理达显著水平(P < 0.05);20天时高盐处理显著高于对照(P < 0.05),低、中盐处理则略低于对照; 30天时,各处理均显著高于对照(P < 0.05)(图 6)。各处理茶条槭在胁迫30天时GSH含量较对照分别增加87.9%,124.9%,96.9%。

|
图 6 不同时间、不同基质NaHCO3浓度胁迫处理下朝鲜接骨木和茶条槭苗木叶片的GSH含量 Figure 6 GSH content in S. coreana and A. ginnala seedlings leaves under different concentrations of NaHCO3 salt stress for different days |
随盐处理浓度的增加,朝鲜接骨木叶片的脯氨酸含量呈现明显上升趋势(图 7)。与对照相比,胁迫10天时,各处理分别增加27.2%(P>0.05),78.2%(P < 0.01),96.4%(P < 0.01);胁迫20天时,各处理分别增加57.6%(P < 0.01),133.0%(P < 0.01),147.9%(P < 0.01);胁迫30天时,各处理分别增加46.4%(P>0.05),51.9%(P < 0.05),72.8%(P < 0.05)。胁迫后期,中、高盐处理脯氨酸含量有所下降。各胁迫处理下茶条槭叶片的脯氨酸含量变化趋势各异(图 7),胁迫10天时,低盐处理显著低于对照,中、高盐处理显著高于对照(P < 0.05);胁迫20天时,各处理呈下降趋势,中盐处理与对照相比差异显著(P < 0.05);胁迫30天时,各处理呈缓慢上升趋势(P>0.05),中盐处理达到最大值。

|
图 7 不同时间、不同基质NaHCO3浓度胁迫处理下朝鲜接骨木和茶条槭苗木叶片的脯氨酸含量 Figure 7 Proline content in S. coreana and A. ginnala seedlings leaves under different concentrations of NaHCO3 salt stress for different days |
随盐处理浓度的增加,2树种可溶性糖含量均呈上升趋势(图 8)。与对照相比,朝鲜接骨木胁迫10天时,各处理分别增加18.8%(P < 0.05),17.3%(P < 0.05),19.4%(P < 0.05);胁迫20天时,各处理分别增加5.1%(P>0.05),7.1%(P>0.05),24.1%(P < 0.05);胁迫30天时,各处理分别增加46.7%(P < 0.05),38.3%(P < 0.05),51.9%(P < 0.05)。与对照相比,茶条槭在胁迫10天时,各处理分别增加18.0%(P < 0.05),23.7%(P < 0.05),49.9%(P < 0.05);胁迫20天时,低盐处理增加3.8%(P>0.05),中盐处理略低于对照(P>0.05),高盐处理增加27.8%(P < 0.05);胁迫30天时各处理分别增加15.6%(P < 0.05), 31.8%(P < 0.05),22.7%(P < 0.05)。随着胁迫时间的延长,2树种的可溶性糖则呈下降趋势,其中茶条槭下降幅度要高于朝鲜接骨木。

|
图 8 不同时间、不同基质NaHCO3浓度胁迫处理下朝鲜接骨木和茶条槭苗木叶片的可溶性糖含量 Figure 8 Soluble sugar content in S. coreana and A. ginnala seedlings leaves under different concentrations of NaHCO3 salt stress for different days |
本研究通过胁迫症状、苗木存活率和苗高、地径生长量3个形态指标的调查来反映2树种苗木在盐胁迫条件下的受害程度。结果显示:胁迫后2树种苗木的存活率在低盐处理下仍为100%,在中、高盐处理下存活率均明显下降(表 1,2),说明它们对低盐胁迫具有较强的适应性,中、高盐胁迫严重影响2树种的生长。本研究中盐胁迫对2树种苗木个体形态的发育产生显著的影响,整体表现是盐胁迫抑制苗木组织和器官的生长。随着盐胁迫的增强,生长随之明显减慢。2树种苗木在低盐处理下受到的抑制程度较小,说明2树种对低盐胁迫具有一定的适应性。综合比较:低盐处理的盐胁迫对朝鲜接骨木和茶条槭的生长抑制作用较小,2树种苗木可以适应低盐处理的盐胁迫。中、高盐处理对朝鲜接骨木和茶条槭的生长抑制作用较大,2树种苗木在中、高盐处理的盐胁迫下不能正常生长。朝鲜接骨木较茶条槭表现的耐盐性稍强。
本研究中,2树种苗木叶片的MDA含量大幅增加(图 1),说明在盐胁迫的过程中,过多的氧自由基引起的膜脂过氧化充当重要角色。MDA从膜上释放后,可以与蛋白质、核酸反应,改变这些大分子的构型,或使之产生交联反应,从而丧失功能,还可使纤维素分子间的桥键松驰,或抑制蛋白质的合成。因此,MDA的积累可能对膜和细胞造成一定的伤害,所以其含量的变化可以反映出细胞膜的受损程度(姜英淑等,2009)。2树种苗木中除MDA含量的大幅上升外,盐胁迫也同样导致其电导率的大幅上升(图 2),电导率的升高是盐胁迫损伤细胞膜的表现, 同时也反映盐敏感植物受盐胁迫后体内的抗氧化系统不足以有效清除这些氧化损伤。同样的报道也见于其他一些植株(Mittova et al., 2002; Hernandez et al., 2000; Sreenivasulu et al., 2000)。在本试验的整个处理过程中,低盐处理时,MDA含量保持较平缓的趋势,表明植株启动自身防御系统时,提高一些酶的活性,可以增强膜脂伤害物质的代谢反应,从而改善叶片的生理代谢功能,提高其膜脂抗过氧化的能力。
本试验中,2树种苗木在盐胁迫下SOD活性呈动态变化(图 3),说明2树种苗木对盐胁迫均产生一定的适应性,胁迫30天后,朝鲜接骨木SOD活性显著降低说明此时朝鲜接骨木清除O2·能力下降,导致膜系统受到严重损害,胁迫30天后,茶条槭的低盐处理略有升高,说明茶条槭对低盐处理浓度的盐胁迫有一定的适应性。朝鲜接骨木CAT在胁迫期间起到一定的保护作用。茶条槭CAT在胁迫前20天起到一定的保护作用(图 4)。盐胁迫期间2树种苗木ASA和GSH含量均增加(图 5,6),这是2树种苗木对盐胁迫的一种适应。
植物体中的各种抗氧化酶和抗氧化剂不是彼此孤立的,而是相互联系,相互作用,彼此构成1个复杂的抗氧化网络,各种酶相互协调,最大限度地清除活性氧,降低活性氧的损伤,提高植物的抗氧化能力,从而提高植物的耐盐能力,使其在盐渍条件下仍能较好地生长发育(Liang et al., 2003; Badawi et al., 2004)。本研究中,盐胁迫的不同梯度和不同时期2树种苗木的抗氧化酶系统在清除活性氧过程中起相互协助作用。
胁迫发生时,植物细胞积累可溶性物质以防止水分流失,使细胞恢复膨胀,应对渗透胁迫(陈少裕,1991; 何承坤等,1996)。本试验认为随盐胁迫浓度的增加,朝鲜接骨木迅速积累脯氨酸含量来维持渗透平衡,从而通过调节抵御盐分对其产生的伤害。茶条槭脯氨酸含量在胁迫10天时,中、高盐处理显著升高,这也是其自身进行积极渗透调节的表现(图 7)。胁迫20天后,各处理脯氨酸积累不明显,说明此时盐胁迫对茶条槭产生一定的伤害。多项研究表明:植物在盐分胁迫下植物体内脯氨酸大量积累(王刚,2009; 韩金龙等,2010)。傅秀云等(1988)研究,不同冬小麦品种耐盐性与脯氨酸含量关系的研究表明,耐盐性强的小麦品种具有较高的脯氨酸含量。但也有不同报道,孙金月等(1997)认为,脯氨酸累积并不代表抗盐能力大小,不能作为抗盐生理指标。关于脯氨酸含量与植物抗盐性关系目前看法还不一致,需作进一步的研究,目前只能将其作为1种辅助指标慎重使用。本试验中,随着盐处理浓度增加,2树种苗木可溶性糖含量均显著升高(图 8),说明可溶性糖对2树种苗木抵御盐胁迫均起到渗透调节的作用。本文的研究结果支持以往的研究结果,即随含盐量的升高,植物叶片可溶性糖含量增加(陈新红等,2008; 田晓艳等,2008; 孙晓光等,2009)。
综上所述,NaHCO3盐胁迫对朝鲜接骨木和茶条槭苗木的生长均产生一定的影响,低盐处理对2树种苗木的生长和生理影响较小,中、高盐处理对2树种的生长和生理产生严重的抑制作用,并且相同胁迫条件下,朝鲜接骨木对碳酸盐的抗性高于茶条槭。朝鲜接骨木和茶条槭苗木对低盐胁迫具有一定的抗性,可耐受的土壤NaHCO3的含量为1.0 g·kg-1。
陈建勋, 王晓峰. 2002. 植物生理学实验指导[M]. 广州: 华南理工大学出版社.
|
陈少裕. 1991. 膜脂过氧化对植物细胞的伤害[J]. 植物生理学通讯, 27(2): 84-90. |
陈新红, 叶裥沣, 周青, 等. 2008. 盐胁迫对小麦幼苗形态和生理特性的影响[J]. 安徽农业科学, 36(33): 14408-14410. DOI:10.3969/j.issn.0517-6611.2008.33.023 |
陈有民. 1990. 园林树木学[M]. 北京: 中国林业出版社.
|
傅秀云. 1988. 冬小麦耐盐力与脯氨酸含量的关系[J]. 山东农业科学, (2): 5-7. |
韩金龙, 徐立华, 徐相波, 等. 2010. 盐胁迫下不同玉米品种在苗期叶片和根中Na+、K+、Ca2+及脯氨酸含量变化的研究[J]. 作物杂志, (1): 49-52. |
何承坤, 郭素枝, 李家慎. 1996. 干旱胁迫对番茄活性氧代谢的影响[J]. 福建农业大学学报, 25(3): 307-311. |
姜英淑, 陈书明, 王秋玉, 等. 2009. 干旱胁迫对2个欧李种源生理特征的影响[J]. 林业科学, 45(6): 6-10. DOI:10.11707/j.1001-7488.20090602 |
李国雷. 2004. 盐胁迫下13个树种反应特性的研究. 山东农业大学硕士学位论文. http://cdmd.cnki.com.cn/article/cdmd-10434-2004121185.htm
|
李合生, 孙群, 赵世杰, 等. 2000. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社.
|
潘保原, 宫伟光, 张子峰, 等. 2006. 大庆苏打盐渍土壤的分类与评价[J]. 东北林业大学学报, 34(2): 57-59. |
孙金月, 赵玉田. 1997. 小麦细胞壁糖蛋白的耐盐性保护作用与机制研究[J]. 中国农业科学, 30(4): 9-12. |
孙晓光, 何青云, 李长青, 等. 2009. 混合盐胁迫下马铃薯渗透调节物质含量的变化[J]. 中国马铃薯, 23(3): 129-132. |
田晓艳, 刘延吉, 郭迎春. 2008. 盐胁迫对NHC牧草Na+、K+、Pro、可溶性糖及可溶性蛋白的影响[J]. 草业科学, (10): 34-38. |
王刚. 2009. 荒漠植物红砂在干旱胁迫和盐胁迫下的渗透调节研究. 兰州大学博士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-10730-2009180611.htm
|
张弼弘, 刘宝光, 刘颖, 等. 2003. 接骨木的开发及利用[J]. 林业科技, 28(5): 53-54. |
赵可夫. 1993. 植物抗盐生理[M]. 北京: 中国科学技术出版社: 18-22, 159, 187-196.
|
Badawi G H, Yamauchi Y, Shimada E, et al. 2004. Enhanced tolerance to salt stress and water deficit by overexpressing superoxide dismutase in tobacco(Nicotiana tabacum)chloroplasts[J]. Plant Sci, 166(4): 919-928. DOI:10.1016/j.plantsci.2003.12.007 |
Hernandez J A, Jimenez A, Mullineaux P, et al. 2000. Tolerance of pea(Pisum sativum L.) to long-term salt stress is associated with induction of antioxidant defenses[J]. Plant and Cell Environment, 23(8): 853-862. DOI:10.1046/j.1365-3040.2000.00602.x |
Liang Y, Chen Q, Liu Q, et al. 2003. Exogenous silicon(Si)increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley(Hordeum vulgare L.)[J]. Plant Physiol, 160(10): 1157-1164. DOI:10.1078/0176-1617-01065 |
Mittova V, Tal M, Volokita M, et al. 2002. Salt stress in duces up-regulation of an efficient chloroplast antioxidant system in the salttolerant wild tomato species Lycopersicon pennellii but not in the cultivated species[J]. Plant Physiol, 115(3): 393-400. DOI:10.1034/j.1399-3054.2002.1150309.x |
Sreenivasulu N, Grimm B, Wobus U, et al. 2000. Differential response of antioxidant compounds to salinity stress in salttolerant and salt-sensitive seedlings of foxtail millet(Setaria italica)[J]. Physiologia Plantarum, 109(4): 435-442. DOI:10.1034/j.1399-3054.2000.100410.x |
Utriainen J, Holopainen T. 2001. Influence of nitrogen and phosphorus availability and ozone stress on Norway spruce seedlings[J]. Tree Physiol, 21(7): 447-456. DOI:10.1093/treephys/21.7.447 |