 2010, Vol. 46
2010, Vol. 46文章信息
- 杨秀艳, 张守攻, 孙晓梅, 宋丛文, 陈柏如, 侯义梅
- Yang Xiuyan, Zhang Shougong, Sun Xiaomei, Song Congwen, Chen Bairu, Hou Yimei
- 北亚热带高山区日本落叶松自由授粉家系遗传测定与二代优树选择
- Genetic Test of Open-Pollinated Larix kaempferi Families and Selection for the Second Generation Elite Trees in Northern Sub-Tropical Alpine Area
- 林业科学, 2010, 46(8): 45-50.
- Scientia Silvae Sinicae, 2010, 46(8): 45-50.
-
文章历史
- 收稿日期:2009-08-04
- 修回日期:2010-01-21
-
作者相关文章



2. 湖北省林业科学研究院 武汉 430075;
3. 湖北省建始县林业局 建始 445300
2. Hubei Academy of Forestry Wuhan 430075;
3. Jianshi Forestry Bureau of Hubei Province Jianshi 445300
日本落叶松(Larix kaempferi)在我国引种有近100年历史,其干形通直,早期速生,材质优良,现已成为我国重要的建筑和纸浆用材树种,主要栽培区由原来的辽东山区扩大到包括四川、湖北、湖南在内的11个省(区)。作为短周期纸浆原料林树种,日本落叶松在中北亚热带山区实现林纸一体化的进程中有着广阔的市场前景。日本落叶松在该区主要引种于海拔1 000~2 100 m的中高山地,在海拔上不与杉木(Cunninghamia lanceolata)和马尾松(Pinus massoniana)等其他用材树种争地,形成该生态区段上造林树种的补充。目前该区已成为我国南方日本落叶松培育基地,栽培面积超过33.3万hm2。基于优树选择具有明显的生态区域性考虑,课题组从20世纪80年代先后在落叶松新引种区广泛营造育种基因资源林和子代测定林,为营造适应该生态区需求的种子园提供了遗传资源保障。同时,以部分子代测定林为材料开展了日本落叶松自由授粉家系子代生长性状、形质性状与材性性状遗传变异及多性状联合选择的研究(孙晓梅等,2003; 2004a;2004b;2005)和营林技术的研究(孙晓梅等,2007a; 2007b;祁万宜等,2007),为该区域日本落叶松遗传改良与栽培利用提供理论指导。
一直以来,北亚热带高山区发展日本落叶松的种苗以东北调拨为主,由于落叶松种子园大小年现象严重,适合该区的种苗更为缺乏,良种成为制约日本落叶松迅速发展的瓶颈,积极营建区域化的高世代种子园已成为发展日本落叶松的迫切需要。遗传基础广泛且品质优良的育种群体是开展高世代育种和组建高世代种子园的关键。二代育种群体的精确评价与选择直接影响到高世代改良水平及高世代种子园的产量与遗传品质。由于表型数据含有环境效应,仅以表型数据很难选择出遗传品质优良的改良材料,育种值剔除了环境影响,反映了真实的遗传效应,提高了选择的精确性。本文以建立在湖北长岭岗国有林场日本落叶松自由授粉家系子代测定林为基础,对生长性状的遗传变异进行评价,并采用最佳线性预测(BLP)方法计算家系及家系内单株的育种值,选出优良家系及二代优树,不仅为加速现有初级种子园的去劣疏伐提供依据,还为高世代改良提供高质量的育种材料,提升当地日本落叶松造林的良种化水平及良种使用率。
1 材料与方法 1.1 试验林概况试验林位于湖北建始长岭岗林场,海拔在1 500 m以上,属北亚热带高山区域。试验地土壤为石灰岩发育而成的山地棕壤,土层深度1 m以上,表土疏松,pH值5~6。从1988年开始,连续5年在林场内共营造了5片日本落叶松自由授粉家系子代测定林。试验采用随机完全区组设计,4~5次重复,6株双列小区。
子代测定林家系种子来自内蒙古旺业甸种子园、山东天麻种子园、辽宁大孤家种子园和湖北长岭岗林场自选优树,具体情况见表 1。5块子代测定林共包含家系119个,其中于1989和1990年营造的子代测定林中有共同家系16个。2006年冬季全面调查各子代测定林的胸径及树高性状,共调查单株1 962株。需要说明的是,由于Ⅱ号子代测定林于1999年进行了间伐,该子代测定林的遗传参数采用1998年的调查数据进行估算。
|
|

材积采用湖北宜昌地区日本落叶松二元材积公式计算: V=0.000 051 082 956 89D1.857 298 121H1. 017 901 505。式中, D,H分别代表胸径和树高。
1.2 研究方法1.2.1统计分析与遗传参数估计性状方差分析采用混合线性模型: Yijk=u+Bi+Fj+FBij+Eijk。式中,Yijk为第i重复第j家系第k单株的观测值,u为总体平均值; Bi为第i重复的固定效应; Fj为第j家系的随机效应,E(Fj)=0,Var(Fj)=σf2; FBij为第i重复第j家系的互作效应,E(FBij)=0,Var(FBij)= σfb2; Eijk为小区内的随机误差,E(Eijk)=0,Var(Eijk)=σe2。由于各家系均有不同程度的缺株,方差分析采用SAS GLM过程计算,方差分量采用PROC VARCOMP过程中的限制最大似然法(REML)计算。
性状的家系遗传力和单株遗传力分别采用以下公式计算(Falconer,1989): hf2=δf2/(δe2/bn+δfb2/b+δf2); hi2=4δf2/(δe2 +δfb2+δf2)。式中,hf2为家系遗传力; hi2为单株遗传力; δf2,δfb2, δe2分别为家系、区组×家系和环境的方差分量; b和n分别表示重复数和小区调和株数。
遗传变异系数和表型变异系数采用下列公式计算: 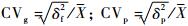
家系选择遗传增益计算公式(黄少伟等,2001)为: ΔG(%)=(x-X)h2/X×100。式中,x为入选家系性状平均值,h2为性状遗传力。
配合选择单株遗传增益的计算公式(周志春等,1999)为: 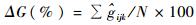
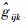
育种值的预测采用最佳线性预测(Best Linear Prediction,BLP)方法(White et al., 1989)。计算公式为: 
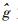
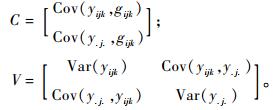
|
以上统计分析利用SAS8.0软件包完成,矩阵运算借助Matlab5.3软件包完成。
2 结果与分析 2.1 生长性状的遗传变异分析对5块子代测定林胸径、树高及材积的方差分析结果(表 2)表明,3个生长性状在家系间均存在显著差异(仅1991年造林的子代家系的树高性状变异幅度较小,但也达到了0.1显著水平),说明各性状在家系间存在丰富的变异。3性状中,变异幅度最大的为材积,遗传变异系数(CVg)和表型变异系数(CVp)分别为12.48%~28.10%,51.16%~63.29%;树高性状变动幅度最小,CVg和CVp分别为2.94%~9.16%,14.15%~22.07%;胸径性状居中,CVg和CVp为4.72%~10.78%。3个性状在5块子代测定林中的遗传力也存在很大的差异。其中1988年造林的子代测定林3性状的家系和单株遗传力最低,在0.5以下; 1989年造林的子代测定林各性状的家系和单株遗传力较高,除树高单株遗传力外,其他均达到0.6以上。除了1988年子代林外,其他4片子代林的家系遗传力均在0.5以上,说明生长性状受较强的遗传控制,开展家系选择能够取得较好的效果。研究表明,3性状的家系遗传力高于单株遗传力,因此二代优树选择应考虑在家系选择基础上进行配合选择。
|
|
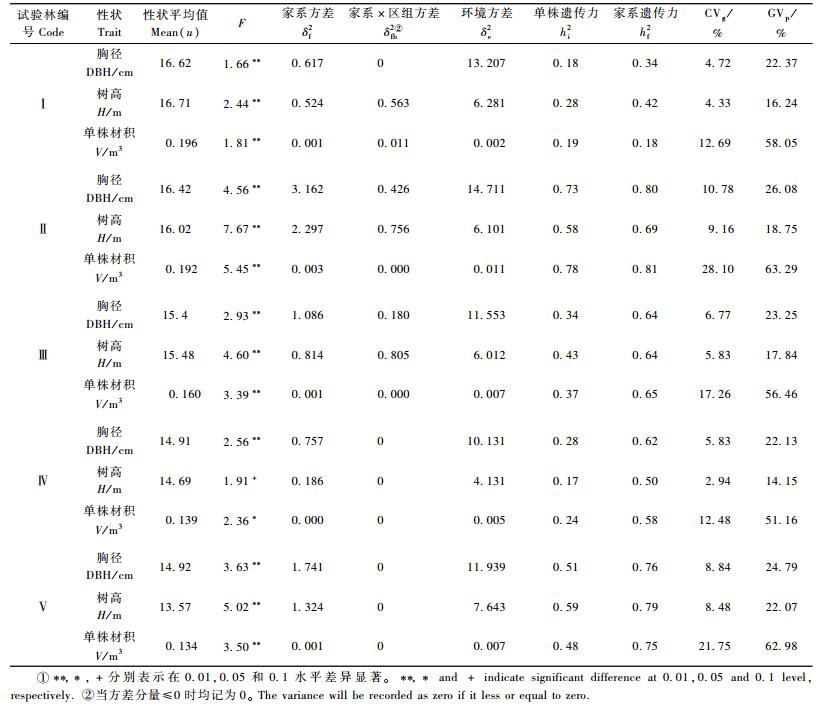
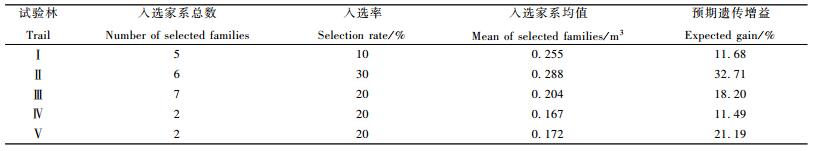
由前面遗传变异分析可以看出,各测定林多数生长性状在家系间的差异达到了显著水平,性状的家系遗传力也普遍较高,因此进行优良家系选择对于生长性状的改良将取得比较理想的效果。以家系材积育种值为选择指标,依据各子代测定林可能获得的期望遗传增益的高低且总的遗传增益不低于10%设定不同的入选率进行家系选择,结果见表 3。由于有的试验林未设遗传对照,因此所有林分均以全部家系的平均值为选择基准计算遗传增益。从表 3可以看出,性状遗传力高且在家系间遗传变异幅度大的遗传增益越大,家系选择的效果越好。
|
|
以子代测定林为基础的选择主要有家系选择、家系内选择及配合选择。在进行二代优树选择时不仅要考虑单株的表现,还要考虑其所在家系同胞的遗传表现。家系与个体的联合选择主要通过2种方式实现:一是把家系平均值与家系内个体值联合计算每个个体的选择指数,并以指数高低进行评选(配合选择); 另一种方式是先选出优良家系,再从入选家系中选出最优个体作为二代优树(家系/家系内选择)。下面对2种方式的选择效果进行比较。
2.3.1 配合选择由White等(1989)给出的BLP计算公式可以看出,预测单株育种值时单株表型值与家系均值各占有一定权重,因此表型值相同的2个单株育种值会因所属家系的不同而异,单株育种值随着家系育种值的升高而增加。根据单株树木的材积测定值与家系平均值,利用BLP方法计算了1 962个单株的育种值。为了尽可能保持较为广泛的遗传多样性,避免近交的过快发生,同时保证较高的遗传增益,最终依据单株育种值的大小和同一子代测定林中同一家系入选优树总数不超过5株的原则(马常耕,1994),以每块子代林总株数的5%~10%的入选比例,共选出优良单株137株,分别来自于49个不同家系(平均每家系入选2.8株); 单株入选率为6.98%,家系入选率为47.12%。
2.3.2 家系/家系内选择优良家系的评定依据家系的育种值和简单t检验(何昭珩等,2001)在不同试验林分别进行评价,对每块林分选出育种值得分显著(t值大于P < 0.05临界值)高于对照(林分总平均值或遗传对照值)的家系,并从入选家系中选择最优的3个单株作为二代优树。根据材积计算结果5块子代测定林共有43个不同家系入选,家系入选率为42.31%,家系/家系内选择入选家系比配合选择减少,2种选择方式入选的家系重叠度为78.3%。入选单株129株,入选率为6.57%,与配合选择相比基本持平。家系/家系内选择所获得的期望遗传增益在第Ⅰ至Ⅳ号子代测定林中与配合选择所获得的增益相比持平或降低,仅在第Ⅴ片子代测定林中比配合选择略高(家系及家系内选择下,单株材积、胸径及树高的期望遗传增益比配合选择分别高6.7%,1.6%和0.6%,表 4)。但在这片子代林中,家系/家系内选择仅有2个家系的6个单株入选,而配合选择则有3个家系的9个单株入选。
|
|
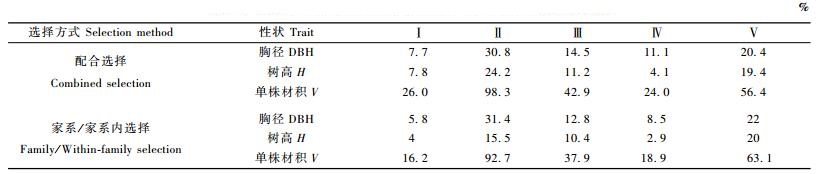
通过对2种选择方式的比较认为,配合选择能够充分兼顾个体及所属家系的遗传表现,所得到的优树群体从所包含家系及单株数量的角度来看,其遗传基础比家系/家系内选择更广泛(将采用分子标记的手段做进一步的验证)。虽然后者有可能得到较高的遗传增益(表 4),但可能要以牺牲遗传多样性为代价,因此建议采用配合选择方式开展二代优树选择。
3 结论与讨论估计育种值的精度是遗传评定的关键环节。常用的育种值估算方法有算术平均数法、小区均值法、最小平方估计等,这些方法均要求数据具有良好的平衡性,但林业调查数据往往不能满足这一要求,造成估算精度不高。最佳线性预测(BLP)在分析复杂、缺失严重的极端不平衡数据方面具有很强的优势。自20世纪80年代末,该方法已成功用于湿地松(Pinus elliottii)(De Souda et al., 1992)、美国短叶松(P. banksiana)(Klein,1995)、马尾松(周志春等,1999)、泡桐属(Paulownia)(马浩等,2003)、小干松(P. contorta ssp. latifolia)(Xie et al., 2007)等树种的育种值预测。BLP方法具有无偏预测、预测值误差方差最小、预测值与真实值相关系数最大等性质,可对不同候选对象求出不同的指数系数,因此可以较好地解决试验中各家系方差不同的问题,是估算日本落叶松家系及单株育种值的理想方法。
本文对日本落叶松自由授粉家系生长性状遗传变异分析发现,日本落叶松自由授粉家系胸径、树高及材积等性状在家系间存在丰富的遗传变异,且各性状的单株遗传力低于家系平均遗传力,因此二代优树的选择在家系选择的基础上进行。通过对配合选择和家系/家系内个体2种选择方式的比较认为,在期望遗传增益相近的情况下,配合选择有更多的家系及个体入选,其遗传多样性可能更为丰富。因此,采用BLP方法预测单株育种值,通过配合选择选出来自于49个家系的二代优树137株,家系和单株入选率分别为47.12%和6.98%。5块子代测定林入选二代优树的材积期望遗传增益分别为26%,98.3%,42.9%,24%和56.4%。
在组建种子园时允许在不同的亚组中存在有亲缘关系的个体,但对优树的亲缘关系应加以控制。有研究认为(Xie et al., 2007)应用具有一定共祖关系的个体建立种子园并不会因近交发生而降低遗传增益,利用完全无亲缘关系的材料建园将使预期增益明显减少。美国南方松改良协作组认为进入下代育种群体中的有亲缘关系的优树最多为5株(马常耕,1994),本研究设定了类似限制原则,并将利用分子标记手段对入选个体进行进一步的亲缘关系筛选。目前,正利用选出的这批二代优树及在北亚热带地区其他试验基地选出的二代优树构建区域化育种群体,建立可控微型育种园,通过树体控制,营养、花期调节等手段缩短结实周期,开展人工交配及子代测定,加速世代转换,并为无性快繁提供采穗母株; 同时将以这批生长优良的二代优树为选择的基础群体,结合物候、开花特性及木材特性研究及分子标记辅助育种手段,进一步筛选出花期一致、结实量大、材质优良、遗传基础广的优树30~60个作为生产群体,通过大砧或高枝嫁接方式营建产量和遗传品质双优的高世代种子园,创新早结实、丰产和稳产的各类管理技术。
何昭珩, 钟伟华. 2001. 应用水平法与t检验评选火炬松半同胞家系[J]. 华南农业大学学报, 22(1): 9-12. DOI:10.7671/j.issn.1001-411X.2001.01.004 |
黄少伟, 钟伟华, 黄凯, 等. 2001. 火炬松自由授粉子代多地点试验[J]. 林业科学研究, 14(5): 509-514. |
马浩, 邓华平, 张冬梅, 等. 2003. 泡桐属植物育种值预测方法的研究[J]. 林业科学, 39(1): 75-80. DOI:10.11707/j.1001-7488.20030112 |
马常耕. 1994. 高世代种子园营建研究的进展[J]. 世界林业研究, (1): 32-38. |
祁万宜, 孙晓梅, 张守攻, 等. 2007. 北亚热带高山区日本落叶松纸浆用材林初植密度的研究[J]. 华中农业大学学报, 26(4): 552-556. |
孙晓梅, 张守攻, 齐力旺, 等. 2003. 日本落叶松自由授粉家系纸浆材材性遗传变异的研究[J]. 林业科学研究, 16(5): 515-522. |
孙晓梅, 张守攻, 王卫东, 等. 2004a. 日本落叶松自由授粉家系形质性状遗传变异的研究[J]. 北京林业大学学报, 26(3): 41-45. |
孙晓梅, 张守攻, 侯义梅, 等. 2004b. 短轮伐期日本落叶松家系生长性状遗传参数的变化[J]. 林业科学, 40(6): 68-74. |
孙晓梅, 张守攻, 李时元, 等. 2005. 日本落叶松纸浆材优良家系多性状联合选择[J]. 林业科学, 41(4): 48-54. DOI:10.11707/j.1001-7488.20050409 |
孙晓梅, 张守攻, 祁万宜, 等. 2007a. 北亚热带高山区日本落叶松幼龄林施肥技术的研究[J]. 林业科学研究, 20(1): 68-73. |
孙晓梅, 张守攻, 祁万宜, 等. 2007b. 北亚热带高山区日本落叶松造林整地与抚育技术的研究[J]. 林业科学研究, 20(2): 235-240. |
周志春, 林荣联, 兰永兆, 等. 1999. 马尾松实生种子园的遗传分析和育种值预测[J]. 林业科学研究, 12(2): 132-138. |
De Souda S M, Hodge G R, White T L. 1992. Indirect prediction of breeding values for fusiform resistance of slash pine parents using greenhouse test[J]. Forest Sci, 38(1): 45-60. |
Falconer D S. 1989. Introduction to quantitative genetics[M]. New York: Longman Scientific & Technical: 438.
|
Klein J I. 1995. Multiple-trait combined seletion in Jack pine family-test plantations using best linear prediction[J]. Silvae Genetica, 44(5/6): 362-375. |
White T L, Hodge G R. 1989.Best linear prediction of breeding values in forest tree improvement. Dordrecht, Netherlands: Kluwer Academic Publishers, 71-84;173-206.
|
Xie C Y, Carlson M R, Murphy J C. 2007. Predicting individual breeding values and making forward selections from open-pollinated progeny test trials for seed orchard establishment of interior Lodgepole pine (Pinus contorta ssp.latifolia) in British Columbia[J]. New Forests, 33(2): 125-138. |