 2010, Vol. 46
2010, Vol. 46文章信息
- 李晓娜, 国庆喜, 王兴昌, 郑海富
- Li Xiaona, Guo Qingxi, Wang Xingchang, Zheng Haifu
- 东北天然次生林下木树种生物量的相对生长
- Allometry of Understory Tree Species in a Natural Secondary Forest in Northeast China
- 林业科学, 2010, 46(8): 22-32.
- Scientia Silvae Sinicae, 2010, 46(8): 22-32.
-
文章历史
- 收稿日期:2009-05-07
- 修回日期:2010-06-06
-
作者相关文章



异速生长广泛存在于植物中(韩文轩等, 2008; Enquist et al., 2002),植物的个体大小与其他属性的异速生长关系并不唯一(Darveau et al., 2002; Jacob, 2004),一般表现为2/3, 3/4和8/3等幂指数关系(韩文轩等, 2008; Enquist et al., 2002; Niklas, 2004),如McMahon等(1976)认为植物高度与茎基部粗度成2/3的幂指数关系以及植物干生物量与茎基部粗度成8/3的幂指数关系。
植物生物量是构成生态系统生产力的重要组成部分,是生态系统运行的物质和能量基础。对生物量测定是深入研究许多林业和生态问题的基础,主要方法有收获法和相对生长法。其中,收获法虽准确但费时费力且不具有连续性,采用相对生长法建立经验回归模型估算植物生物量有效地降低对植被的破坏,更重要的是可以得到植物生物量的连续变化。国内外已建立很多森林类型乔木生物量的相对生长模型,并整理了特定树种和具体立地的经验相对生长方程,用于森林生态系统生物量和生产力的研究(冯宗炜等, 1999; Crow, 1978; Nelson et al., 1999; Sah et al., 2004; Wang, 2006)。同时,国外研究者建立不少灌木生物量相对生长模型,用于准确估算森林碳储量及退化森林生态系统的恢复等研究(Alaback, 1987; Brown, 1976; Paton et al., 2002),国内一些学者采用相对生长法也构建了生物量模型用于干旱半干旱地区灌丛生物量(姜凤岐等,1982; 陈遐林等,2002)以及南方人工林下木层植物生物量(曾慧卿等, 2007)的估算等,但大多灌木生物量模型很少考虑不同树种和器官最佳生物量模型的差异(张峰等,1993)。东北地区仅潘攀等(2007)根据长白落叶松(Larix olgensis)人工林上层乔木的林龄和林分密度等因子估算了其灌木层的生物量,基于植物自身属性的温带天然次生林下木树种生物量模型的研究尚未见报道。
林下植物生物量仅占森林总生物量中较小的部分,但其植被化学物质含量和生物量归还率比上层植物还高,对养分循环的作用不可低估,在维护整个森林生态系统的演替发展、功能稳定和生物多样性等方面起着重要作用(杨昆等, 2006),东北天然次生林下木层发育良好,在水源涵养、水土保持、生态系统的演替发展等方面起着重要作用(陈大珂等, 1990)。本研究以帽儿山天然次生林下木树种为研究对象,构建植物生物量的相对生长方程,分析各器官生物量与植株大小的相对生长关系,为准确估算温带次生林森林生物量和生产力,评价温带天然次生林下木层的生态功能提供科学依据和基础数据。
1 材料与方法 1.1 研究地概况试验地位于黑龙江省帽儿山森林生态系统国家野外科学观测站(127°30′—127°34′ E,45°20′—45°25′ N),该地区植被属于长白植物区系,是东北东部山区典型的天然次生林区。该区天然次生林大致分为硬阔林、白桦林、山杨林和蒙古栎林4种林型。林区内的乔木树种主要有春榆(Ulmus davidiana var. japonica)、色木槭(Acer mono)、胡桃楸(Juglans mandshurica)、白桦(Betula platyphylla)、水曲柳(Fraxinus mandschurica)、山杨(Populus davidiana)、黄菠萝(Phellodendron amurense)、紫椴(Tilia amurensis)和蒙古栎(Quercus mongolica)等。林下植物发育良好,主要树种有暴马丁香(Syringa reticulata var. amurensis)、茶条槭(Acer ginnala)、稠李(Padus asiatica)、兴安鼠李(Rhamnus davuricus)、东北鼠李(Rhamnus yoshinoi)、疣点卫矛(Euonymus verrucosoides)、卫矛(Euonymus alatus)、珍珠梅(Sorbaria sorbifolia)、乌苏里绣线菊(Spiraea ussuriensis)、东北山梅花(Philadelphus schrenkii)、金银忍冬(Lonicera maackii)、鸡树条荚蒾(Viburnum sargentii)、暖木条荚蒾(Viburnum burejaeticum)、东北溲疏(Deutzia parviflora var. amurensis)、刺五加(Acanthopanax senticosus)和毛榛(Corylus mandshurica)等。
1.2 数据获取选取东北东部天然次生林下木层主要树种(陈大珂等, 1990)作为研究对象(表 1),依据植物形态,将其分为典型灌木和乔木型植物2类(Sah et al., 2004),即珍珠梅、乌苏里绣线菊、东北山梅花、忍冬、东北溲疏、暖木条荚蒾和刺五加等地面分枝多主干不明显的7种植物为典型灌木,暴马丁香、茶条槭、稠李、兴安鼠李、东北鼠李、疣点卫矛、卫矛和毛榛等地面分枝少、主干明显的8个树种为乔木型植物,鸡树条荚蒾地面多个主干,暂将其列为乔木型植物。2008年8—9月在帽儿山森林生态站取样,依据植物生长状况,在各植物的核心分布地段选取一定数量的植株全收获,测量树高(H,m)、冠幅(假设冠幅为椭圆形,CA=π×Cl×Cw/4,长轴Cl,m; 短轴Cw,m)、基径(D0,mm)、离地10 cm处树干直径(D10,mm)、分叉高(Hca,m)、分叉径(Dca,mm)和胸径(DBH,mm),将植物整株挖出,深度为根系分布所达范围,测量主干长(L,m),分叶、新枝、多年枝、细根和粗根,称其鲜质量(精确到1 g),将所有称量的组分各取适量样品放入85 ℃恒温干燥箱内烘至恒量, 计算干物质含量及各器官干质量。各植物个体生物量干质量和易测因子的分布范围见表 1。
|
|

选取与植物各器官生物量叶(ML)、新枝(MNB)、多年枝(MOB)、细根(MSR)、粗根(MLR)、总枝(MBR)、地上部分(MAG)、地下部分(MBG)以及个体生物量(MT)相关性显著(P < 0.05)的植物属性作为自变量,分别采用逐步回归法和选用线性方程y=a+bx、二次多项式y=a+bx+cx2、三次多项式y=a+bx+cx2+dx3、对数方程y=a+blnx、幂函数y=axb和指数方程y=aebx等6种函数建立单物种(单一物种器官生物量与易测因子的相对生长方程)和混合物种(多个物种器官生物量与易测因子的相对生长方程)2类相对生长方程。依据方程的显著性以及R2,FI,SE和CF来评价方程的优劣,选出方程显著(P < 0.05)、标准误较小、拟合性较好的数学模型作为最佳模型来估算生物量,比较分析单物种模型与混合物种模型的优劣以及自变量中分别引入树高和主干长的生长方程与仅基于树干直径(D)或冠幅(CA)等生物量方程的优劣。所有统计分析均用SPSS 13.0完成。
比较经对数转换的乘幂方程和未经对数转换的方程时,选用标准误修正因子CF (Sprugel, 1983)和适合指数FI (Brand et al., 1985)替代乘幂方程的标准误SE和决定系数R2作为模型优劣的判断指标,当乘幂方程的FI值与线性方程、二次多项式或三次多项式的R2接近时,选择乘幂方程作为最优模型。CF和FI值的计算公式如下:
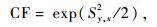
|
(1) |
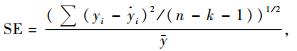
|
(2) |

|
(3) |
式(1)中,y是因变量,x是自变量,Sy, x是y=axb的标准误(SE),其计算公式见式(2)。式(2)和(3)中yi是第i个植株生物量实测值,y是植株生物量的平均值,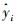
分析植物叶、新枝、多年枝、细根和粗根等器官以及总枝、地上部分、地下部分和个体的生物量与个体大小的相对生长关系,即乔木型植物生物量与树干直径的相对生长关系(M∝D)和典型灌木生物量与冠幅(CA)或冠高乘积(CAH)的相对生长关系(M∝CA或M∝CAH)。相对生长规律通常用幂函数的形式表示(韩文轩等,2008),取对数后即:
lgy=lgβ+αlgx,
式中y为植物器官生物量,x为植物属性,α为器官生物量与植物属性的幂指数,β为生长常数。理论上,α与器官种类和植物属性特征有关,而与植物种类和环境等无关,β随植物种类或环境的变化而变化。
2 结果与分析 2.1 各植物相对生长方程下木植物器官生物量的幂函数模型和最佳估算方程大多显著(P < 0.05)(仅鸡树条荚蒾的新枝生物量方程的显著性为0.52),R2值大多超过0.800,2类生长型植物的最佳生物量模型的方程形式均是以幂函数为主,包括少数线性方程和二次多项式。
2.1.1 乔木型植物单物种生物量模型暴马丁香、稠李、茶条槭、疣点卫矛、卫矛、毛榛、东北鼠李、兴安鼠李和鸡树条荚蒾9种乔木型植物器官生物量均用D10解释较理想(表 2),各器官生物量最佳估算模型的自变量大多为D10,只有兴安鼠李新枝的最佳生物量模型基于D102H。大多植物器官最佳生物量模型的形式为幂函数,其中毛榛的叶和新枝的最佳生物量模型的形式为线性方程,茶条槭和疣点卫矛的粗根和地下部分、稠李的叶和新枝的最佳生物量模型的形式为二次多项式。
|
|
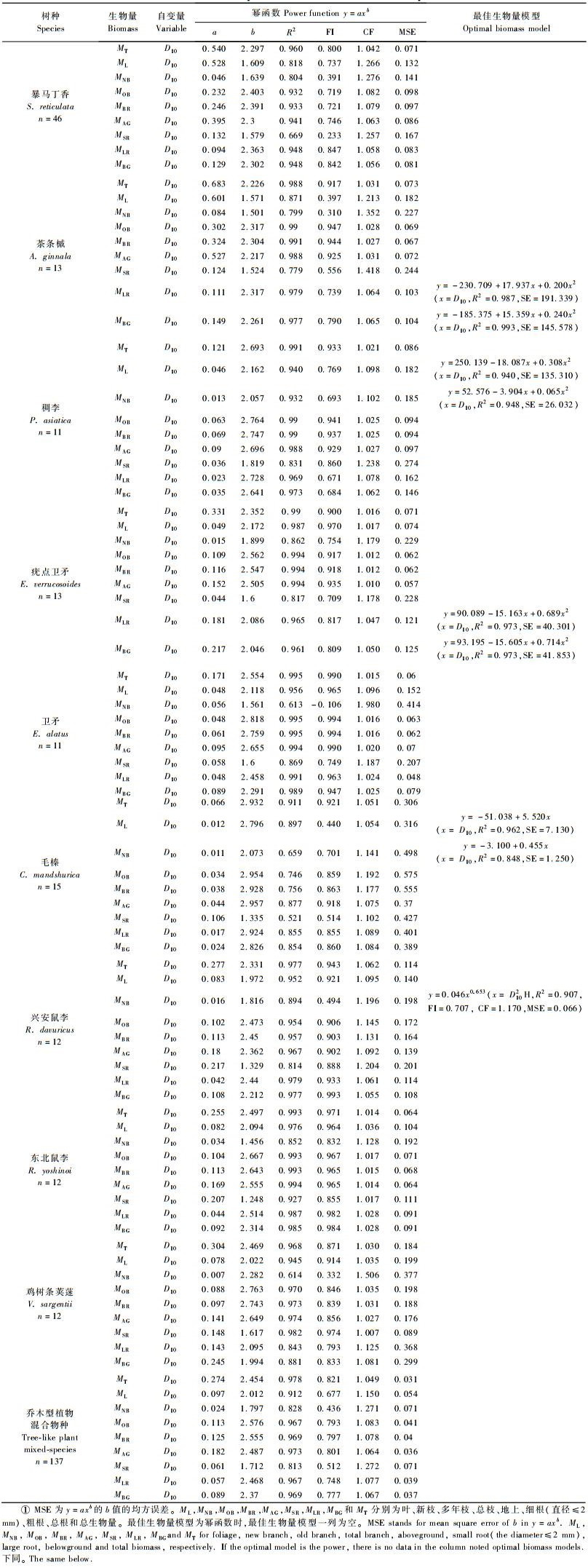
不同植物各器官生物量的最佳估算方程均有较大的R2值和较小的SE值(表 2)。其中总枝、地上部分和地下部分以及个体生物量生长方程的R2值的范围分别是0.756(毛榛)~0.995(卫矛)、0.877(毛榛)~0.994(疣点卫矛)、0.854(毛榛)~0.993(茶条槭)、0.911(毛榛)~0.995(卫矛),叶、新枝、多年枝、粗根和细根等器官生物量生长方程的R2值的变化范围依次是0.818(暴马丁香)~0.987(疣点卫矛)、0.613(卫矛)~0.993(鸡树条荚蒾)、0.746(毛榛)~0.995(卫矛)、0.843(鸡树条荚蒾)~0.991(卫矛)和0.521(毛榛)~0.982(鸡树条荚蒾)。整体上,各器官生物量最佳估算方程的R2值相比,从大到小依次是多年枝>粗根>叶>新枝>细根; 各树种生物量最佳生长方程的R2值的大小依次是东北鼠李>稠李>疣点卫矛>兴安鼠李>茶条槭>卫矛>鸡树条荚蒾>暴马丁香>毛榛。
2.1.2 典型灌木单物种生物量模型典型灌木器官生物量均用冠幅CA或冠高乘积CAH等自变量解释较好(表 3),如乌苏里绣线菊、暖木条荚蒾、刺五加和东北溲疏各器官以及东北珍珠梅的叶、多年枝、总枝和地上生物量、东北山梅花的叶和新枝等器官的生物量用CAH解释较好,忍冬各器官和珍珠梅的新枝、地下部分和个体生物量以CA为自变量估算较理想,仅东北山梅花大多器官(叶和新枝除外)生物量最佳预测变量为树高。各植物器官最佳生物量模型的方程形式多为幂函数,少数是线性方程和二次多项式,如刺五加的新枝、东北溲疏的新枝和东北山梅花的粗根和地下部分的最佳方程为线性方程,忍冬和乌苏里绣线菊的新枝、暖木条荚蒾的细根、粗根和地下部分等器官生物量的最佳方程是二次多项式。
|
|
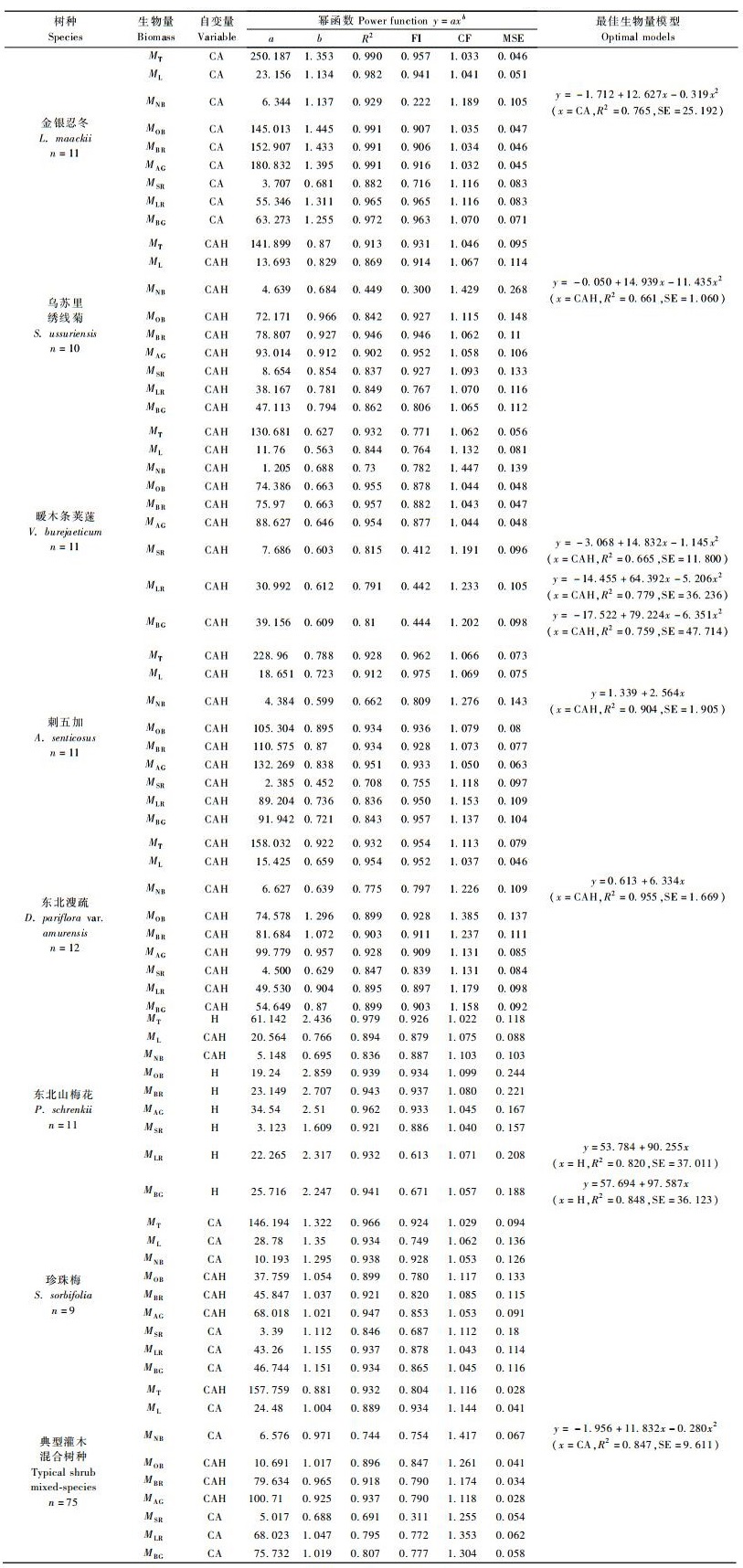
典型灌木各器官最佳生物量模型的拟合性均较好(表 3)。总枝、地上部分和地下部分以及个体最佳生物量模型的R2值的范围分别是0.903(东北溲疏)~0.991(金银忍冬)、0.902(乌苏里绣线菊)~0.991(金银忍冬)、0.759(暖木条荚蒾)~0.972(金银忍冬)、0.913(乌苏里绣线菊)~0.990(金银忍冬),叶、新枝、多年枝、粗根和细根等器官最佳生物量模型的R2值的范围依次是0.844(暖木条荚蒾)~0.982(金银忍冬)、0.661(乌苏里绣线菊)~0.955(东北溲疏)、0.842(乌苏里绣线菊)~0.991(金银忍冬)、0.665(暖木条荚蒾)~0.921(东北山梅花)和0.779(暖木条荚蒾)~0.965(金银忍冬)。
与乔木型植物生物量模型相比,典型灌木器官生物量最佳估算模型的R2值稍低,各器官最佳生长方程的R2值从大到小依次是多年枝>叶>粗根>新枝>细根,各树种生物量估算模型的优劣依次是金银忍冬>珍珠梅>东北溲疏>东北山梅花>刺五加>乌苏里绣线菊>暖木条荚蒾。
2.1.3 混合物种生物量模型本研究中2类混合物种最佳生物量模型(7种典型灌木的混合物种模型和9种乔木型植物的混合物种模型)的方程形式均以幂函数为主,仅典型灌木新枝生物量的生长方程为二次多项式,但2类混合物种生物量模型的自变量和拟合性均有差异(表 2,3)。乔木型植物器官生物量均用D10解释较好,但对于典型灌木来说,叶、新枝、细根、粗根和地下部分等器官生物量用CA解释较理想,而多年枝、总枝、地上部分和个体的生物量用CAH估算较准确。
2类混合物种大多数器官(典型灌木的新枝和细根除外)最佳生物量模型的R2值虽多低于单物种模型,但大多也在0.800以上(表 2, 3)。乔木型植物器官生物量混合物种最佳模型的R2值在0.813(细根)~0.978(个体)之间,与其相比,暴马丁香各器官, 毛榛的多数器官(叶、新枝和细根除外), 兴安鼠李的多年枝、总枝、地上部分和个体, 茶条槭的叶、新枝和细根, 卫矛的新枝, 鸡树条荚蒾的粗根和地下部分和个体等生物量的单物种最佳模型的R2值稍低。典型灌木生物量混合物种最佳模型的R2值在0.691(细根)~0.932(个体)之间,稍高于乌苏里绣线菊的叶、新枝、多年枝、地上部分和个体, 暖木条荚蒾的叶、新枝、细根、粗根和地下部分, 东北溲疏的总枝和地上部分, 金银忍冬和东北山梅花的新枝以及刺五加个体生物量的单物种模型。
整体上,乔木型植物中毛榛(0.814±0.129)、暴马丁香(0.884±0.099)和鸡树条荚蒾(0.906±0.119)3种植物的单物种生物量模型的R2值稍低于混合物种模型(0.931±0.066),典型灌木中暖木条荚蒾(0.830±0.115)和乌苏里绣线菊(0.846±0.083)的单物种模型的R2值稍低于混合物种模型(0.863±0.084)。可见,多数树种和器官的单物种生物量模型优于混合物种模型。
2.1.4 自变量中引入树高(H)和主干长(L)对生物量方程的贡献与基于D10(CA)单变量生物量模型的R2值相比,引入H变量和L变量的生物量模型的R2值的变化趋势大致相同,但其变化幅度因树种和器官而异(图 1)。

|
图 1 树高和主干长对生物量模型的决定系数(R2)的贡献 Figure 1 Percentage increase in determination coefficient (R2) if adding tree height and stem length into the D10-only or CA-only biomass equations a为引入树高和主干长后各树种生物量模型的R2值的变化,b为引入H和L后两类生长型植物器官生物量模型的R2值的变化。a中树种标识同表 1,TOT, FOL, BR, AG和BG分别是植物的个体、叶、总枝、地上部分和地下部分。 a is the percentage increase in determination coefficient (R2) if adding tree height or stem length into the D10-only or CA-only biomass equations for understory, b is the percentage increase in determination coefficient (R2) if adding tree height or stem length for component biomass equations of two growth type plants, a by species noted as the same as in Tab. 1, and b by biomass components. TOT, FOL, BR, AG and BG stand for the biomass of individual, foliage, total branch, aboveground and belowground, respectively. |
引入树高后生物量模型的R2值的变化因树种和器官而异(图 1)。典型灌木生物量模型的R2值的增加量的范围在-1.36%±1.54%(珍珠梅)~15.27%±3.11%(东北山梅花)之间,只有珍珠梅生物量模型的R2值减小; 乔木型植物生物量模型的R2值变化的范围是-1.54%±1.76%(鸡树条荚蒾)~1.62%±1.31%(毛榛),其中暴马丁香(0.48%±0.82%)、茶条槭(0.10%±0.26%)、稠李(0.04%±0.41%)和毛榛4种植物的R2值增加(图 1)。
2类生长型植物各器官生物量模型的R2值的变化不同(图 1)。乔木型植物器官生物量模型的R2值的增加量在-0.92%±1.39%(叶生物量模型)~0.23%±1.30%(枝生物量模型),其中枝和地上部分的生物量模型的R2值增加(图 1); 典型灌木器官生物量模型的R2值的增加量的范围是6.99%±6.73%(个体生物量)~3.56%±5.30%(叶生物量)(图 1)。可见,树高能够有效提高典型多数灌木器官生物量模型的R2值,而对于多数乔木型植物来说,用D10就能较好地解释其器官生物量。
与引入树高后生物量模型的R2值的变化相比,主干长L对各植物器官生物量模型的R2值的贡献较小(图 1)。引入主干长后各植物生物量模型的R2值变化的范围是-9.28%±5.02%(珍珠梅)~13.40%±1.79%(东北山梅花),乔木型植物中稠李(0.24%±0.09%)、疣点卫矛(0.08%±0.15%)和毛榛(1.12%+1.49%)3种植物的生物量模型的R2值增加(图 1); 典型灌木生物量模型的R2值的变化与引入树高后相似,变化范围在-9.28%±5.02%(珍珠梅)~13.40%±1.79%(东北山梅花),只有珍珠梅生物量模型的R2值减小。2类生长型植物各器官生物量模型的R2值的变化与引入树高后相似(图 1),乔木型植物器官生物量模型的R2值的变化范围是-0.89%±1.28%(叶)~0.36%±1.20%(枝),其中枝和地上部分的R2值增加,典型灌木各器官生物量模型的R2值的变化在2.70%±9.49%(地下部分)~4.86%±8.40%(个体)。
2.2 各树种器官生物量与易测因子的相对生长关系乔木型植物器官生物量与D10的相对生长关系以及典型灌木器官生物量与冠幅CA或冠高乘积CAH的相对生长关系均极显著(P < 0.001),但相对生长指数并不唯一。
2.2.1 乔木型植物器官生物量与D10的相对生长关系乔木型树种器官生物量与D10的相对生长关系在1.712~2.555,叶、新枝、多年枝、细根、粗根、总枝、地上部分、地下部分和个体的生物量与D10的相对生长关系在95%置信区间CI(下同)分别是1.906~2.119, 1.658~1.937, 2.496~2.657, 1.572~1.852, 2.390~2.546, 2.477~2.633, 2.416~2.558, 2.298~2.443和2.392~2.517,其中多年枝、粗根、总枝、地上部分、地下部分和个体的生物量与D10的相对生长关系接近理论值8/3。
2.2.2 典型灌木器官生物量与冠幅CA和冠幅乘以高度CAH的相对生长关系典型灌木器官生物量与冠幅的相对生长关系在0.688~1.293,叶、新枝、多年枝、细根和粗根各器官生物量以及总枝、地上部分、地下部分和个体的生物量与CA的相对生长关系分别是1.004(0.921~1.086,R2=0.889), 0.971(0.838~1.104,R2=0.744), 1.293(1.144~1.443,R2=0.803), 0.688(0.580~0.795,R2=0.691), 1.047(0.923~1.171,R2=0.795), 1.236(1.107~1.364,R2=0.834), 1.192(1.081~1.304,R2=0.861), 1.019(0.902~1.135, R2=0.807)和1.133(1.024~1.243, R2=0.853),其中叶、新枝、粗根、地下部分和个体的生物量与CA近于等速生长,多年枝、总枝和地上部分的生物量与冠幅的相对生长指数接近4/3,细根与冠幅的幂函数比例近于0.67。
各器官生物量与CAH的相对生长关系在0.436~1.017之间,叶、新枝、多年枝、细根和粗根各器官生物量以及总枝、地上部分、地下部分和个体的生物量与CAH的相对生长关系分别是0.761(0.709~0.812,R2=0.922), 0.717(0.616~0.818,R2=0.733), 1.017(0.936~1.097,R2=0.896), 0.527(0.452~0.601,R2=0.731), 0.816(0.743~0.889,R2=0.872), 0.965(0.897~1.032,R2=0.918), 0.925(0.869~0.982,R2=0.937), 0.793(0.726~0.860,R2=0.884), 0.881(0.826~0.937,R2=0.932),其中多年枝、总枝和地上部分与CAH近于等速生长,叶和地下部分的生物量与CAH的相对生长关系近于0.75,新枝生物量与CAH的相对生长关系介于0.67~0.75,粗根生物量与CAH的幂函数比例介于0.75~0.86,个体生物量与CAH的相对生长关系接近0.86,只有细根生物量与CAH的幂函数比例不足0.6。
3 结论与讨论 3.1 下木树种的生物量模型植物生物量模型自变量的选择与自身形态关系密切(Paton et al., 2002)。一些灌木树种的最佳生物量模型以枝茎直径作为自变量(陈遐林等, 2002; Alaback, 1986; Paton et al., 2002),有些灌木以植株盖度或冠幅为自变量估算生物量较好(曾慧卿等, 2007; Halpern et al., 1996)。有研究表明地面分枝多的丛生灌木的枝茎直径与各器官生物量的相关系数大于冠幅,但用枝茎直径作为自变量的回归模型预测生物量误差较大(张峰等, 1993; Brown, 1976),Brown(1976)用基径构建方程估算灌木生物量,但建议对于分枝多植株矮小的灌木用盖度和高度作为自变量。
与大乔木不同,林下植物生物量模型中增加第2个变量(如树高H或主干长L),整体上并不能使模型的R2值大幅上升(图 1)。本地区下木植物生物量模型中仅用D10就能较好地解释乔木型植物的器官生物量,而增加H能够有效提高典型灌木生物量模型的拟合性,表明植株形态接近圆柱形的植物的生长不仅表现在冠幅的横向生长还包括高度的纵向伸长(Halpern et al., 1996)。引入H变量和L变量对生物量模型的R2值的贡献相似(图 1),大多典型灌木(东北山梅花除外)和少数乔木型植物的生物量模型的R2值提高。与H相比,引入L变量的生物量模型的R2值增加较小。
本地区下木树种最佳生物量估算模型的方程形式多以幂函数为主(表 2, 3),其FI值并不一定小于R2值(Sah et al 2004),仅少数植物的新枝、细根和粗根等个别器官的最佳生物量模型的方程形式为二次多项式或线性方程,这与曾慧卿等(2007)的研究结果有差别,曾慧卿等建立的红壤丘陵区林下灌木生物量估算模型的方程形式多为二次多项式。
本地区下木树种生物量模型的R2值与他人报道的生物量模型的R2值的变异范围相一致(姜凤岐等, 1982; 陈遐林等, 2002; 曾慧卿等, 2007; Paton et al., 2002)。下木树种各器官生物量模型的拟合性相比均是细根和新枝的模型最差,乔木型植物最佳生物量模型的R2值普遍优于典型灌木,表明生物量模型的准确度可能与物种生长形态有关。通常认为,单物种生物量模型优于混合物种模型(Buech et al., 1989; Sah et al., 2004),尤其是叶、小枝和细根等物质转换周期短的器官(表 2,3)(Buech et al., 1989),本研究中并不是所有的单物种模型都优于混合物种模型,对于尚未建立单物种生物量模型的植物,可以选择混合物种模型或植株形态特征相似的单物种模型来估算其生物量(Brown, 1976; Crow, 1978; Nelson et al., 1999)。
3.2 下木植物生物量的相对生长规律温带天然次生林下木植物生物量与其他属性的相对生长关系均显著(P < 0.05)(韩文轩等, 2008; Enquist et al 2002; McMahon et al., 1976; Niklas, 2004),相对生长指数并不唯一,具有可塑性(Darveau, 2002; Jacob, 2004)。乔木型植物器官生物量与D10的相对生长关系在1.712~2.555,其中多年枝、粗根、总枝、地上部分、地下部分和个体生物量与D10的幂函数比例接近理论值8/3(McMahon et al., 1976)。典型灌木器官生物量与冠幅的相对生长关系在0.688~1.293,其中叶、新枝、粗根和地下部分的生物量与冠幅近于等速生长; 器官生物量与冠高乘积的相对生长关系在0.527~1.017,其中多年枝、总枝和地上部分的生物量与冠高乘积的相对生长关系近于1。
3.3 生物量模型的适用性相对生长关系给高大树木的生长量或固碳能力的测定提供了理论基础和便利条件(Enquist et al., 2002; Niklas, 2004)。本研究的对象为下木树种,与大乔木树种相比(Wang, 2006),直径变化范围较小。然而,本地区下木发育良好,很多树种属于小乔木或大灌木。这些树种的空间异质性较大,如果用研究草本和小灌木常用的全收获法来估算其生产力,尽管在理论上讲得通,但实际应用上存在两大问题:一是空间异质性可能导致随机设置的样方失去代表性,二是收获法费时费力,对于暴马丁香、稠李、茶条槭和兴安鼠李等乔木型树种来说难以实现(曾慧卿等, 2007; Sah et al., 2004)。然而,对于个体较小且地上分枝较多恢复较快的典型灌木而言,收获法对样地的干扰相对较小,往往比测树法更为实用(Paton et al 2002)。因此建议,如果研究目的是估算下木生物量,采用收获法或测树法(相对生长法)或者二者结合,主要考虑破坏性是否为关键因素; 如果研究生产力,对乔木型树种来说,宜采用测树法,而典型灌木宜采用收获法。
本研究完善了我国温带森林的主要树种生物量生长方程数据库,同时为普遍相对生长理论增加了基础数据。然而,森林下木层物种的生长状况因其立地条件存在一定的差异,可能出现同一物种在不同立地条件下的最佳估算生物量模型不同。本研究针对温带天然次生林下木植物,在研究区外应用本文建立的生物量估算模型时需进行验证; 本地区下木植物自身属性如基径、D10、冠幅等超过本研究中下木植物自身属性的分布范围时,使用本研究中的生物量方程估算生物量时也需做验证。
陈大珂, 周晓峰, 祝宁, 等. 1994. 天然次生林——结构·功能·动态与经营[M]. 哈尔滨: 东北林业大学出版社: 53-57.
|
陈遐林, 马钦彦, 康峰峰, 等. 2002. 山西太岳山典型灌木林生物量及生产力研究[J]. 林业科学研究, 15(3): 304-309. |
冯宗炜, 王效科, 吴刚. 1999. 中国森林生态系统的生物量和生产力[M]. 北京: 科学出版社: 8-12.
|
韩文轩, 方静云. 2008. 幂指数异速生长机制模型综述[J]. 植物生态学报, 32(4): 951-960. |
姜凤岐, 卢风勇. 1982. 小叶锦鸡儿灌丛地上生物量的预测模型[J]. 生态学报, 2(2): 103-110. |
罗云建, 张小全, 王效科, 等. 2009. 森林生物量的估算方法及其研究进展[J]. 林业科学, 45(8): 129-134. DOI:10.11707/j.1001-7488.20090823 |
潘攀, 牟长城, 孙志虎. 2007. 长白落叶松人工林灌丛生物量的调查分析[J]. 东北林业大学学报, 35(4): 1-6. |
杨昆, 管东生. 2006. 林下植被的生物量分布特征及其作用[J]. 生态学杂志, 25(10): 1252-1256. DOI:10.3321/j.issn:1000-4890.2006.10.019 |
张峰, 上官铁梁, 李素珍. 1993. 关于灌木生物量建模方法的改进[J]. 生态学杂志, 12(3): 67-69. |
曾慧卿, 刘琪璟, 冯宗炜, 等. 2007. 红壤丘陵区林下灌木生物量估算模型的建立及其应用[J]. 应用生态学报, 18(10): 2185-2190. |
Alaback P B. 1986. Biomass regression equations for understory plants in coastal Alaska: Effects of species and sampling design on estimates[J]. Northwest Science, 60: 90-103. |
Brown J K. 1976. Estimating shrub biomass from basal stem diameters[J]. Canadian Journal of Forest Research, 6: 154-158. |
Buech R R, Rugg D J. 1989. Biomass relations of shrub components and their generality[J]. Forest Ecology and Management, 26: 257-264. DOI:10.1016/0378-1127(89)90086-8 |
Crow T R. 1978. Biomass and production in contiguous forests in northern Wisconsin[J]. Ecology, 59: 265-273. DOI:10.2307/1936372 |
Darveau C A, Suarez R K, Andrews R D, et al. 2002. Allometric cascade as a unifying principle of body mass effects on metabolism[J]. Nature, 417: 166-170. DOI:10.1038/417166a |
Enquist B J, Niklas K J. 2002. Global allocation rules for patterns of biomass partitioning in seed plants[J]. Science, 295: 1517-1520. DOI:10.1126/science.1066360 |
Gehring C, Park S, Denich M. 2004. Liana allometric biomass equations for Amazonian primary and secondary forest[J]. Forest Ecology and Management, 195: 69-83. DOI:10.1016/j.foreco.2004.02.054 |
Halpern C B, Miller E A, Geyer M A. 1996. Equations for predicting aboveground biomass of plant species in early successional forests of the western Cascade Range, Oregon[J]. Northwest Science, 70: 306-320. |
Jacob W. 2004. Allocation, plasticity and allometry in plants[J]. Perspectives in Plant Ecology, Evolution and Systematics, 6: 207-215. DOI:10.1078/1433-8319-00083 |
McMahon T A, Kronauer R E. 1976. Tree structures: deducing the principle of mechanical design[J]. Journal of Theoretical Biology, 59: 443-466. DOI:10.1016/0022-5193(76)90182-X |
Murray R B, Jacobson M Q. 1982. An evaluation of dimension analysis for predicting shrub biomass[J]. Journal of Range Management, 35: 451-454. DOI:10.2307/3898603 |
Nelson B W, Mesquita R, Pereira J L G, et al. 1999. Allometric regressions for improved estimate of secondary forest biomass in the central Amazon[J]. Forest Ecology and Management, 117: 149-167. DOI:10.1016/S0378-1127(98)00475-7 |
Niklas K J. 2004. Plant allometry: is there a grand unifying theory?[J]. Biological Reviews, 79: 871-889. DOI:10.1017/S1464793104006499 |
Paton D, Nuńez J, Bao D, et al. 2002. Forage biomass of 22 shrub species from Monfragüe Natural Park (SW Spain) assessed by log-log regression models[J]. Journal of Arid Environments, 52: 223-231. DOI:10.1006/jare.2001.0993 |
Sah J P, Ross M S, Koptur S, et al. 2004. Estimating aboveground biomass of broadleaved woody plants in the understory of Florida Keys pine forests[J]. Forest Ecology and Management, 203: 319-329. DOI:10.1016/j.foreco.2004.07.059 |
Sprugel D G. 1983. Correcting for bias in log-transformed allometric equations[J]. Ecology, 64: 209-210. DOI:10.2307/1937343 |
Wang C. 2006. Biomass allometric equations for 10 cooccurring tree species in Chinese temperate forests[J]. Forest Ecology and Management, 222: 9-16. DOI:10.1016/j.foreco.2005.10.074 |