 2010, Vol. 46
2010, Vol. 46文章信息
- 张启昌, 张英楠, 其其格
- Zhang Qichang, Zhang Yingnan, Qi Qige
- 长白山阔叶红松林的红松种群热值
- Caloric Values of Pinus koraiensis Population in Broad-Leaved Korean Pine Forests in Changbai Mountain
- 林业科学, 2010, 46(8): 15-21.
- Scientia Silvae Sinicae, 2010, 46(8): 15-21.
-
文章历史
- 收稿日期:2009-03-30
- 修回日期:2010-04-09
-
作者相关文章


2. 北京林业大学林学院 北京 100083;
3. 吉林省林业调查规划院 长春 130022
2. Forestry College of Beijing Forestry University Beijing 100083;
3. Jilin Academy of Forest Inventory, Planning Changchun 130022
植物热值是指植物干物质完全燃烧后所释放出来的能量值,它反映绿色植物在光合作用中固定太阳辐射能的能力,是植物本身的重要特征,也是评价和反映生态系统中物质循环和能量转化规律的重要指标。随着生态系统功能过程研究的深入,热值测定日益引起人们的重视(官丽莉等,2005)。自Long(1934)首先开展植物热值研究以来,对生态系统中各种物质的热值及其变化机制的研究日趋广泛(Neitzke,2002; Alexandrov,2008)。国内在这方面的研究虽然起步较晚,但也开展了大量的研究工作(Lin et al., 2008),如对草地(Bao et al., 2007),高寒草原(于应文等,2000),森林(陈美玲等,2009),农田(杨京平等,2002),竹林(林益明等,2000),湿地(李萍萍等,2008)和水生植物(林鹏等,1991)等热值进行了研究。植物不同器官热值直接关系到整株植物能量的分配与贮存,对于了解植物生长特性和生长具有重要的意义。
长白山阔叶红松林主要分布在海拔500~1 100 m的范围内,处于云冷杉针叶林带与阔叶林带之间,是长白山垂直分布带谱中面积最大的一带(李建东等,2001)。长白山阔叶红松林是长白山地区生物种类最丰富的地带性植被,因具有较大的面积和较高的生产力而成为森林碳循环研究的重要对象,被列入全球变化研究的中国东北样带,也是ChinaFLUX的观测站点之一(周广胜等,2002); 长白山阔叶红松林是目前世界上保存较为完好的原始森林类型之一,对于调解径流与气候、维系区域陆地生态平衡有着重要意义,在我国的森林资源和生态环境建设中具有重要的地位和作用。很多学者相继在长白山阔叶红松林的群落特征(外貌特征、种类组成、垂直结构),生物多样性,空间分布格局,净生态系统碳交换,细根分布及其周转,主要树种光合作用、年龄结构与径级结构,林隙,生态价位,生态系统的呼吸速率,降雨截留量的估算,土壤水分物理性质,生态系统土壤呼吸作用,热量平衡变化特征,冠层空气动力学参数等方面做了大量的研究工作(王战等, 1980; 阳含熙等,1985; 郝占庆等,1994; 于振良等,2001; 王蕾等,2009)。但目前还未见有关长白山阔叶红松林红松热值研究的文献报道。研究阔叶红松林红松种群热值的一般规律,可为红松及其所在群落能量特征和生态效率研究提供理论基础和科学依据。本文通过对长白山阔叶红松林不同径级、不同构件、不同季节、不同群落红松热值的研究,探讨有关影响植物热值的因素,为长白山森林生态系统能流研究提供基础资料和科学依据。
1 材料与方法 1.1 研究地自然概况研究地点位于长白山自然保护区内的阔叶红松林带,地理位置为127°33′—128°16′ E,41°42′—42°45′ N,年平均气温在0.9~4.0 ℃,年雨量630~780 mm,海拔500~1 100 m,林分郁闭度0.9,土壤类型为暗棕色森林土。该林分结构为复层混交异龄林,林分组成为3红松(Pinus koraiensis)+3紫椴(Tilia amurensis)+1色木槭(Acer mono)+1蒙古栎(Quercus mongolica)+1水曲柳(Fraxinus mandshurica)+其他,林分密度300~400株·hm-2。蓄积量400~500 m3·hm-2,灌木层盖度0.4,草本层盖度可达0.8(代力民等,2002)。
在研究不同群落热值时,以长白山北坡暗针叶林带中海拔1 200 m处的藓类云冷杉红松林为对照。其群落常见于长白山海拔1 100~1 500 m的地带,面积不大,多成岛状分布于暗针叶林带中。地形多为台地,坡度较小,土壤类型为棕色针叶林土,具有亚寒带气候特点,是以红松(占蓄积的33%)、鱼鳞云杉(Picea jezoensis,18%)、红皮云杉(Picea koraiensis,24%)、臭冷杉(Abies nephrolepis,7%)、长白落叶松(Larix olgensis,15%)和其他(3%)组成的针叶混交林(徐化成,2001)。
1.2 采样野外调查及采样在长白山北坡海拔740 m的典型椴树阔叶红松林和海拔1 200 m的藓类云冷杉红松林内进行。春季(4月)和秋季(10月)各取样1次。在选择样株时,分下木、亚乔木和乔木3个层次7个类型的红松为研究对象。下层木中选取树高30~50 cm(地径小于3 cm)的幼树记为D0;亚乔木层分为2个类型,地径3~4 cm记为DB4,地径5~6 cm记为DB6;乔木层分为4个类型,胸径5~10 cm记为D8,胸径10~15 cm植株(不包括10 cm)记为D13,胸径15~25 cm植株记为D20;胸径25 cm以上植株记为D26,每一径级的样本选取15~20株做重复。分别取叶、枝、干、根和皮,叶和枝分1、2年生叶, 多年生叶,1、2年生枝和多年生枝,根分为主根和侧根。树叶按树冠从上到下分3层,在最下层再区分3层,按比例取样后混合均匀; 树枝考虑东、西、南、北4个方位,按比例取样; 树干部分在距地面1.3 m处用生长锥取样; 树根将主根和侧根按比例混合均匀后再进行取样; 因春季D0苗木较小,未取到其春季皮。在藓类云冷杉红松林群落内选用红松径级分布最多的D20作为不同群落研究的对比径级,采集干、皮、主根和侧根,选取20株做重复。
1.3 测定方法和数据分析采集回来的试验材料首先在105 ℃烘箱下杀青3 h,之后在自然状态下风干1周,再拿到70 ℃烘箱中烘干72 h,再将植物样放入植物粉碎机中粉碎,粉碎后的植物样过0.2 mm筛备用。仪器利用江苏鹤壁天宇仪器有限公司生产的ZDHW-2000微机全自动量热仪。采用量热法测定热值,计算机程序控制。苯甲酸标定量热系统,人工充氧、放气,系统自动测定热值(每样品重复测定5次,取平均值),样品热值以干质量热值表示。
所测数据用SPSS13.0软件进行方差分析(ANOVA),用Tukey法进行多重比较。文中数据以平均值和标准误(Mean±SE)表示。
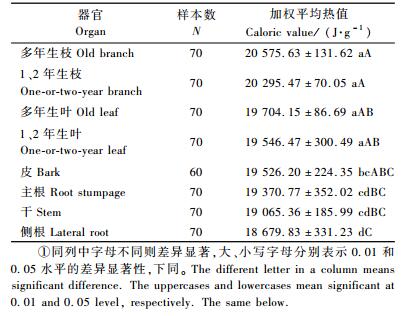
2 结果与分析 2.1 不同器官的热值不同器官对热值的影响达极显著水平(F=5.055,P < 0.01),说明不同器官间的热值差异很大。各器官热值的大小顺序为:多年生枝(20 575.63 J·g-1)>1、2年生枝(20 295.47 J·g-1) >多年生叶(19 704.15 J·g-1) >1、2年生叶(19 546.47 J·g-1)>皮(19 526.20 J·g-1)>主根(19 370.77 J·g-1)>干(19 065.36 J·g-1)>侧根(18 679.83 J·g-1)。从表 1可知,多年生枝, 1、2年生枝, 多年生叶, 1、2年生叶相互之间的差异未达到显著程度(P>0.05),而它们与皮、主根、干、侧根之间的差异显著(P < 0.05)。多年生枝, 1、2年生枝分别与侧根之间的差异极显著(P < 0.01)。从全株植物的平均热值角度分析,枝>叶>皮>干>根,对于枝和叶,多年生的热值要高于1、2年生的热值,对于根来说,主根要高于侧根。
|
|
以春季红松热值为研究对象,对不同径级红松热值的变化规律进行研究。由于红松各器官的生物量不同,因此对各器官加权平均得出红松热值(表 2)。
|
|

不同径级红松的热值差异很大,达到极显著水平(F=4.271,P < 0.01)。通过表 2可以看出,DB4~D20径级之间的差异未达到显著的程度(P>0.05),而D13和DB6与D0差异均极显著(P < 0.01)。红松的热值不是随着径级的增大而增加,它呈明显的波浪形分布,在DB6时达到第1个高峰, 为19 682.30 J·g-1,然后随径级增加呈下降趋势,之后到D13达到最高峰为19 884.84 J·g-1,然后又开始下降,到D26时又出现第3峰为19 050.31 J·g-1。径级30~50 cm的红松幼树热值显著低于其他径级的红松热值(P < 0.05)。
2.3 不同季节的热值从图 1~3中可以看出各径级秋季热值明显高于春季热值,1、2年生叶春季的热值随着径级增大基本持平,而秋季热值则呈明显的单峰型; 多年生叶在春、秋2个季节的热值变化规律基本与1、2年生叶相同。树枝春季的热值随着径级增大而增加,到D20达到最大, 为21 030.33 J·g-1,之后稍有下降,而秋季则呈双峰型即M型,但DB6热值要比D20大。树干春季的热值随着径级增大呈W型,以D13最大为19 956.83 J·g-1,而秋季则呈明显的单峰型,以DB4最大,热值达到22 450.33 J·g-1; 树皮春季的热值随着径级增大呈N型,以DB6最大为20 642.83 J·g-1,而秋季则呈单峰型,以DB6最大,热值达到22 445 J·g-1; 主根春季的热值随着径级增大呈M型,以DB6最大为20 347.17 J·g-1,而秋季则呈明显的单峰型,以DB4最大,热值达到24 094.00 J·g-1; 侧根春季的热值随着径级增大呈W型,以D26最大为19 880.61 J·g-1,而秋季则呈明显的N型,以DB6最大为21 215.17 J·g-1。

|
图 1 不同季节红松各径级叶的热值变化 Figure 1 Seasonal change of caloric values of leaves of P. koraiensis in different diameter-classes |

|
图 2 不同季节红松各径级枝、干和皮的热值变化 Figure 2 Seasonal change of caloric values of branches, stems and barks of P. koraiensis in different diameter-classes |

|
图 3 不同季节红松各径级根的热值变化 Figure 3 Seasonal change of caloric values of roots of P. koraiensis in different diameter-classes |
春季同一径级不同器官热值情况, 对于D0,枝最高,主根最低,分别为19 941.50 J·g-1和15 455.00 J·g-1,按热值高低排序如下:枝>干>多年叶>1、2年叶>侧根>主根; 对于DB4,枝最高,侧根最低,分别为20 282.17 J·g-1和19 200.83 J·g-1,按热值高低排序如下:枝>主根>皮>多年叶>1、2年叶>干>侧根; 对于DB6,皮最高,侧根最低,分别为20 642.83 J·g-1和19 077.67 J·g-1,按热值高低排序如下:皮>枝>主根>多年叶>1、2年叶>干>侧根; 对于D8,枝最高,侧根最低,分别为20 494 J·g-1和17 684.17 J·g-1,按热值高低排序如下:枝>皮>多年叶>1、2年叶>主根>干>侧根; 对于D13,枝最高,1、2年叶最低,分别为20 760.2 J·g-1和19 163.5 J·g-1,按热值高低排序如下:枝>干>多年叶>主根>皮>侧根>1、2年叶; 对于D20,枝最高,侧根最低,分别为21 030.33 J·g-1和17 789.75 J·g-1,按热值高低排序如下:枝>多年叶>1、2年叶>干>皮>主根>侧根; 对于D26,枝最高,干最低,分别为20 360 J·g-1和18 759 J·g-1,按热值高低排序如下:枝>多年叶>主根>侧根>1、2年叶>皮>干。
秋季同一径级不同器官热值情况, 对于D0,枝最高,侧根最低,分别为22 100.00 J·g-1和19 698.50 J·g-1,多年生叶要高于1、2年生叶,按热值高低排序如下:枝>皮>干>多年叶>1、2年叶>主根>侧根; 对于DB4,枝最高,1、2年叶最低,分别为23 849.67 J·g-1和21 215.17 J·g-1,多年生叶要高于1、2年生叶,按热值高低排序如下:枝>多年叶>1、2年叶>干>皮>主根>侧根; 对于DB6,多年叶最高,干最低,分别为25 980.5 J·g-1和20 355 J·g-1,按热值高低排序如下:多年叶>枝>1、2年叶>皮>干>主根>侧根; 对于D8,枝最高,干最低,分别为21 633 J·g-1和20 014.67 J·g-1,按热值高低排序如下:枝>皮>主根>多年叶>侧根>1、2年叶>干; 对于D13,枝最高,干最低,分别为21 050.67 J·g-1和20 122.67 J·g-1,按热值高低排序如下:枝>多年叶>1、2年叶>主根>皮>侧根>干; 对于D20,枝最高,侧根最低,分别为21 728.25 J·g-1和19 146.5 J·g-1,按热值高低排序如下:枝>多年叶>1、2年叶>主根>皮>干>侧根; 对于D26,侧根最高,干最低,分别为20 804.5 J·g-1和19 663.67 J·g-1,按热值高低排序如下:枝>主根>多年叶>侧根>1、2年叶>皮>干。
2.4 不同群落的热值红松在阔叶红松林和暗针叶林带高海拔藓类云冷杉红松林这2个群落都有分布,因此,有必要对比分析这2个群落的红松热值情况。通过图 4可以明显看出,无论干、皮、主根还是侧根,暗针叶林带高海拔藓类云冷杉红松林红松的热值都高于阔叶红松林带红松的热值,以侧根热值差距最大,差值为3 471 J·g-1,热值差距最小的为树干,差值仅为1 075 J·g-1。

|
图 4 红松不同群落热值变化 Figure 4 Caloric values of P. koraiensis in different communities |
本研究红松枝、叶之间差异不显著,而它们与皮、主根、干、侧根之间的差异显著(P < 0.05)。枝与侧根之间的差异极显著(P < 0.01)。红松各器官的平均热值从大到小依次为枝、叶、皮、干和根,对于枝和叶,多年生的热值要高于1、2年生的热值,对于根来说,主根要高于侧根。植物各器官热值的差异可能与其成分和生理功能密切相关。已有的研究表明,植物组分干物质热值的高低,直接受植物体内脂肪、蛋白质和碳水化合物的影响。几种含能物质的热值常数分别为:粗蛋白22.990 kJ·g-1,粗脂肪38.874 kJ·g-1,粗纤维17.556 kJ·g-1(Bliss,1962),无氮浸出物17.135 kJ·g-1(Lieth et al., 1975),碳水化合物17.991 kJ·g-1(Sinclair,1990)。从植物解剖学和植物生理学角度看, 叶是植物体生理活动最活跃的器官,植物进行光合作用的场所,含有较多的高能化合物, 如蛋白质和脂肪等物质; 繁殖体花、果实、种子、胚轴等含有大量高能的粗脂肪和蛋白质; 干、枝和皮是植物体的支持或营养运输器官,组成以纤维素和木质素为主,纤维素和木质素的热值相对蛋白质和脂肪低; 而根部则残留了大量从土壤中吸收的低能矿物质和氧化物,灰分含量高(Bidwell,1982)。因此,多数植物的叶、繁殖体的干质量热值较高,干、枝、皮的热值其次,根最低(陈美玲等,2009; 曾小平等,2009)。但也有一些植物表现出叶的热值低于枝、皮或根的热值,如牛皮桦(Betula utilis)、大叶白颜(Kandelia candel)、山月桂(Gironniera subaequalis)、绿竹(Dendrocalamopsis oldhami)、金露梅(Potentilla fruticosa)表现出枝的热值大于叶的热值(官丽莉等,2005); 山黄麻(Trema orientalis)、云南银柴(Aporusa yunnanensis)等有些短命植物把本身大部分的能量投资到繁殖上,而不是个体生长上,导致它们的叶片具有低热值,表现出枝的热值大于叶的热值(乔秀娟等,2007); 甜槠(Castanopsis eyrei)树枝和树皮的热值大于叶的热值,黄山松(Pinus taiwanensis)树皮的热值大于叶的热值,其原因主要为枝、皮含有较高的高能物质,特别是含有较高的树脂和松节油(林益明等,1999)。红松枝的热值高于叶的热值,其原因是红松枝中含有较多的树脂和松节油。
本研究红松不同径级间的热值差异极显著,树高30~50 cm的红松幼树热值显著低于其他径级的红松热值(P < 0.05)。红松热值在DB6时达到第1个高峰为19 682.30 J·g-1,然后随径级增加呈下降趋势,之后到D13达到最高峰为19 884.84 J·g-1,然后又开始下降,到D26时又出现第3峰为19 050.31 J·g-1。说明红松热值随径级增大呈波浪型分布,在其他树种种群中也有类似现象(倪穗等,2001)。关于植物热值随年龄(径级)的变化规律,胡宝忠等(1998)对白三叶(Trifolium repens)种群的热值测定表明,热值与年龄相关,特别是根的能值与年龄显著相关,年龄越大,能值越高; 刘庆等(1995)对斑苦竹(Pleioblastus maculatus)无性系种群的能量结构研究表明,竹子的年龄对热值有一定影响,斑苦竹无性系分株的平均热值随着年龄级的增加,呈Logistic曲线变化。处于长白山阔叶红松林下层幼苗阶段的红松热值显著低于其他径级的红松热值,说明从幼树到成年过渡阶段,热值随年龄的增加而增加。由于林内环境垂直分异,从乔木层到下木层光照呈指数下降,受太阳辐射越弱则热值降低越多(商侃侃等,2008)。所研究的阔叶红松林的林分郁闭度达到0.9,下层的红松幼树处于乔木层的树荫之下,受太阳光照较弱,体内能量积累有限,高能物质含量相对匮乏,其热值相对要低于乔木层的红松。而处于亚乔木层和乔木层中的红松各径级热值之间虽然呈现波浪型分布,但其差异不显著(P>0.05),说明此阶段红松的径级(年龄)对其热值的影响不大。亚乔木层和乔木层中的红松处于群落的上层,接受的太阳能较多,基本能够满足红松的正常生长,光合作用产生大量的有机物质,积累的高能化合物也相应较多,其热值则较高,且差异不明显。
红松各器官秋季热值均高于春季。这与Singh等(1980)对印度萨瓦纳群落中植被地上部分及阮志平等(2007)对厦门市布迪椰子(Butia capitata)不同器官热值的研究结果一致。植物春季、夏季干质量热值的水平比较低,秋季较高,这种变化与植物的生长规律有一定关系,因为植物在春、夏季植物生长迅速,导致植物体内的能量物质含量受到稀释而减少; 到秋季植物地上部分生长速度慢,但是光合作用仍然很强,体内多糖合成多,同时分配给各组分的光合产物也多,同时由于温度逐渐降低,积累的有机物促使干质量热值的提高以增强抗寒力,所以,植物体内积累较多的能量使得植物体内的能量含量增加到最高值(阮志平等,2007)。但James等(1978)研究发现叶片在春季热值最高。Wielgolaski等(1975)认为植物叶干质量热值的季节变化趋势因不同种类而不同或相似。热值随季节变化与植物的物候节律及对环境因子变化的反应有关,并不是固定不变的(陈美玲等,2009)。由于秋季温度逐渐降低,红松累积有机物质促使其提高热值增加抗寒能力,对高能物质的积累逐渐增加,因此红松秋季热值高于春季。
高海拔藓类云冷杉红松林红松的各器官热值高于阔叶红松林带红松的相应器官热值,以侧根热值差距最大(3 471 J·g-1),干的热值差距最小(1 075 J·g-1)。本文所调查的高海拔藓类云冷杉红松林样地海拔1 200 m,阔叶红松林带样地海拔740 m,二者海拔相差较大,热值呈现为高海拔大于低海拔。对华盛顿高山植物的能量研究证实高山地区植物热值较高(Hadley et al., 1964)。对西双版纳热带季节雨林和高海拔哀牢山中山湿性常绿阔叶林优势植物的热值研究也确认高海拔植物热值大于低海拔的观点(黄钰辉等,2007)。这是由于高海拔的植物种类,因海拔的升高,温度降低,紫外线辐射增强,从而植物的呼吸作用减弱,其叶片组织中酶活性增强, 使体内非结构性碳水化合物积累的能力高于低海拔或温暖地区的植物种类; 高海拔地区太阳光谱成分中短波光所占的比例较高,而短波光中的蓝、紫光有助于植物蛋白质的合成(龙瑞军等,1993)。
陈美玲, 上官周平. 2009. 黄土高原子午岭林区6个典型群落优势种的热值和养分特征[J]. 林业科学, 45(3): 140-144. DOI:10.11707/j.1001-7488.20090324 |
代力民, 孙伟中, 邓红兵, 等. 2002. 长白山北坡椴树阔叶红松林群落主要树种的年龄结构研究[J]. 林业科学, 38(3): 73-77. DOI:10.11707/j.1001-7488.20020314 |
官丽莉, 周小勇, 罗艳. 2005. 我国植物热值研究综述[J]. 生态学杂志, 24(4): 452-457. |
郝占庆, 赵士洞, 陶大立. 1994. 长白山北坡阔叶红松林草本植物物种多样性及其季节动态[J]. 生物多样性, 2(3): 125-132. DOI:10.17520/biods.1994021 |
胡宝忠, 刘娣, 周以良, 等. 1998. 白三叶无性系植物种群分株间的资源分配[J]. 东北林业大学学报, 26(2): 25-28. |
黄钰辉, 官丽莉, 周国逸, 等. 2007. 西双版纳热带季节雨林和哀牢山中山湿性常绿阔叶林优势植物及地表凋落物层的热值[J]. 植物生态学报, 31(3): 457-463. |
李建东, 吴榜华, 盛连喜. 2001. 吉林植被[M]. 长春: 吉林科学技术出版社: 98-99.
|
李萍萍, 陆军, 吴沿友, 等. 2008. 镇江滨江湿地优势植物种群的热值及其能量生产动态[J]. 浙江大学学报:农业与生命科学版, 34(2): 221-229. |
林鹏, 林光辉. 1991. 几种红树植物的热值和灰分含量研究[J]. 植物生态学与地植物学学报, 15(2): 114-120. |
林益明, 林鹏. 1999. 福建武夷山2个典型植物群落建群种的热值研究[J]. 武夷科学, 15: 118-123. |
林益明, 李和阳, 林鹏, 等. 2000. 福建南靖虎伯寮亚热带雨林竹类植物热值的研究[J]. 竹子研究汇刊, 19(1): 57-62. |
刘庆, 钟章成. 1995. 斑苦竹无性系种群能量结构研究[J]. 渝州大学学报:自然科学版, 12(3): 22-27. |
龙瑞军, 徐长林, 胡自治, 等. 1993. 天祝高山草原15种饲用灌木的热值及季节动态[J]. 生态学杂志, 12(5): 13-16. |
倪穗, 陈启瑺. 2001. 青冈种群的热值研究[J]. 浙江大学学报:农业与生命科学版, 27(4): 390-392. |
乔秀娟, 曹敏, 林华. 2007. 西双版纳不同林龄次生植物群落优势树种的热值[J]. 植物生态学报, 31(2): 326-332. DOI:10.17521/cjpe.2007.0038 |
阮志平, 杨志伟, 李元跃, 等. 2007. 布迪椰子不同器官热值的季节变化研究[J]. 热带亚热带植物学报, 15(5): 399-402. |
商侃侃, 陈波, 达良俊. 2008. 浙江天童受损常绿阔叶林实验生态学研究(Ⅲ):从热值角度分析常绿阔叶林常见种的适应策略[J]. 华东师范大学学报:自然科学版, (4): 25-30. |
王蕾, 张春雨, 赵秀海. 2009. 长白山阔叶红松林的空间分布格局[J]. 林业科学, 45(5): 54-59. DOI:10.11707/j.1001-7488.20090508 |
王战, 徐振邦, 李昕, 等. 1980. 长白山北坡主要森林类型及其群落结构特点[J]. 森林生态系统研究, (1): 26-42. |
徐化成. 2001. 中国红松天然林[M]. 北京: 中国林业出版社: 64-65.
|
阳含熙, 王本南, 韩进轩. 1985. 长白山北坡阔叶红松林数量分类[J]. 森林生态系统研究, (5): 15-30. |
杨京平, 博尼菲斯, 姜宁, 等. 2002. 稻田施氮水平对两种水稻热值变化特征影响[J]. 生态学报, 22(2): 240-246. |
于应文, 胡自治, 张德罡. 2000. 天祝金强河高寒地区金露梅的热值及其季节动态[J]. 草业科学, 17(2): 1-4. |
于振良, 于贵瑞, 赵士洞, 等. 2001. 长白山阔叶红松林动态及经营管理模型研究[J]. 资源科学, 23(6): 59-63. |
周广胜, 王玉辉, 蒋延玲. 2002. 全球变化与东北样带(NECT)[J]. 地学前沿, 9(1): 198-216. |
曾小平, 蔡锡安, 赵平, 等. 2009. 广东鹤山人工林群落主要优势植物的热值和灰分含量[J]. 应用生态学报, 20(3): 485-492. |
Alexandrov G A. 2008. Forest growth in the light of the thermodynamic theory of ecological systems[J]. Ecological Modelling, 216(2): 102-106. DOI:10.1016/j.ecolmodel.2007.11.022 |
Bao Y J, Li Z H, Han X G, et al. 2007. Caloric content of plant species and its role in a Leymus chinensis steppe community of Inner Mongolia, China[J]. Acta Ecologica Sinica, 27(11): 4443-4451. DOI:10.1016/S1872-2032(08)60002-5 |
Bidwell R G S. 1982. Plant physiology (Volume Ⅱ)[M]. Beijing: Higher Education Press: 173-181.
|
Bliss L C. 1962. Caloric and lipid content in alpine tundra plants[J]. Ecology, 43(4): 753-757. DOI:10.2307/1933473 |
Hadley E B, Bliss L C. 1964. Energy relationships of alpine plant of Mt[J]. Washington, New Hampshire. Ecological Monographs, 34: 331-357. DOI:10.2307/2937067 |
James T D W, Smith D W. 1978. Seasonal changes in the caloric values of the leaves and twigs of Populus tremuloides.[J]. Can J Bot, 56: 1804-1805. DOI:10.1139/b78-216 |
Lieth H, Whittaker R. 1975. Primary productivity of the biosphere[M]. New York: Springer-Verlag: 119-129.
|
Lin H, Cao M. 2008. Plant energy storage strategy and caloric value[J]. Ecological Modelling, 217(2): 132-138. |
Long F L. 1934. Application of calorimetric methods to ecological research[J]. Plant Physiol, 9(2): 323-327. DOI:10.1104/pp.9.2.323 |
Neitzke M. 2002. Changes in energy fixation and efficiency of energy capture in above-ground biomass along an environmental gradient in calcareous grasslands[J]. Flora, 197: 103-117. DOI:10.1078/0367-2530-00020 |
Sinclair T R. 1990. Nitrogen influence on the physiology of crop yield//Rabbinge R, Goudrin J, Penningg D E, et al. Theoretical production ecology: reflection and prospects. Wageningen: Pudoc, 41-52. http://www.sciencedirect.com/science/article/pii/016819239290127P
|
Singh A K, Misra K N, Ambasht R S. 1980. Energy dynamics in a savanna ecosystem in India[J]. Jap J Ecol, 3: 295-305. |
Wielgolaski F E, Kjelvik S. 1975. Energy content and use of solar radiation of Fennoseandian Tundra plants//Wielgolaski F E. Fennoseandian Tundra ecosystem, Part Ⅰ: Plants and microorganisms. Berlin: Springer Verlag, 201-207. http://link.springer.com/10.1007/978-3-642-80937-8_22
|