 2010, Vol. 46
2010, Vol. 46文章信息
- 张猛, 王丹, 汤浩茹, 任少雄, 刘仁道
- Zhang Meng, Wang Dan, Tang Haoru, Ren Shaoxiong, Liu Rendao
- 费约果茎解剖结构与插条不定根的形成
- Anatomical Structure of Stem and Adventitious Root Formation of Feijoa Cuttings
- 林业科学, 2010, 46(7): 183-187.
- Scientia Silvae Sinicae, 2010, 46(7): 183-187.
-
文章历史
- 收稿日期:2009-02-01
- 修回日期:2010-04-02
-
作者相关文章



2. 四川农业大学林学园艺学院 雅安 625014
2. College of Forestry and Horticulture, Sichuan Agricultural University Ya'an 625014
费约果(Feijoa sellowiana)是桃金娘科多年生亚热带常绿灌木,原产于巴西东南部和乌拉圭,因其果实香味类似凤梨(Ananas comosus),又名凤梨番石榴(Thorp et al., 2002)。费约果在新西兰、美国、法国、西班牙、俄罗斯、澳大利亚、日本等国有栽培,我国江苏、上海、四川、浙江等南方地区也有一定规模的种植。费约果作为一种集观赏、食用与药用为一体的新兴果树,在食品、药品、化妆品等领域具有广阔的应用前景(Nakashima, 2001; Hardy et al., 1970; Kolesnik et al., 1991)。在四川省外国专家局引智项目资助下,2004年,西南科技大学王丹教授从新西兰引进少量费约果苗木后,对其在四川地区的适应性以及综合性状进行了初步研究,认为费约果能够适应四川地区气候,并能保持其优良的食用与观赏特性,是一种极具发展潜力的食用兼观赏的树种(王丹等, 2007)。
费约果繁殖可通过实生繁殖和扦插繁殖2种方式。实生繁殖比较容易,但性状变异较大,难以保持费约果的优良性状,不宜在生产上采用。扦插繁殖能保持原有的性状且简便易行,是生产中广为应用的繁殖技术。然而,在扦插繁殖过程中往往会遇到生根困难的问题(Figueiredo et al., 1995)。如何加快费约果苗木的繁殖速度,从而满足生产对其苗木的需求是当前迫切需要解决的问题。目前对费约果生根困难的原因在解剖学上的研究未见报道,本研究从费约果茎解剖结构与其生根特性的关系和不定根发生、发育的形态解剖学入手,阐述根系在发生发育过程中的变化机制,为探明费约果扦插生根机制和广泛推广费约果的种植奠定理论和技术基础。
1 材料与方法2007年5月20日、6月20日从不同树龄(1,2,3,4年生)费约果采集当年生健壮、无病虫的半木质化枝(semi-hard branch,SB), 10月20日采集木质化枝(hard branch,HB),所采插条于西南科技大学设施农业实验室进行扦插试验。插条长度8~10 cm,保留2~3个节位,上端留两半叶,上切口平剪,下切口(插条形态学下端)距叶柄0.2 cm处斜剪。选用1 000 mg·L-1的IBA作为促根剂速蘸10 s,扦插基质为珍珠岩。采用随机区组试验设计,30枝插条为1个处理, 重复3次。插后40天移栽时,调查各处理的平均生根率。
扦插前留取不同树龄母树的枝条,用以比较研究插条的解剖结构与其生根特性的关系。以6月20日扦插的1年生母树半木质化枝插条为试材,研究费约果插条不定根的发生、发育过程,扦插时随机取5枝插条,以后每隔5天取样1次,直至插条产生不定根。观察插条的外部形态,并从插条基部切取1~2 cm茎段,用FAA固定液固定保存,乙醇-甘油软化1周,采用常规石蜡切片法制片(李正理, 1987),切片厚度10 μm;番红-固绿染色,OLYMPUS-DMB5型显微镜观察并拍照。
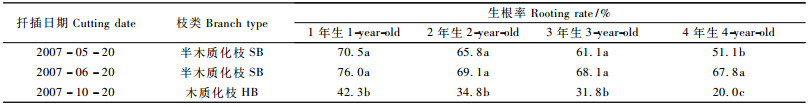
2 结果与分析 2.1 不同扦插时期、不同树龄的费约果插条生根情况不同扦插时期、不同树龄费约果插条的生根结果表明,插条生根具有明显的年龄效应,其生根能力与插条的生理成熟度和扦插时期密切相关。从表 1可以看出,扦插时期对生根率有较大影响,6月20日扦插生根情况最好,以半木质化枝扦插效果优于木质化枝。随着母树年龄增长,半木质化枝和木质化枝插条生根率明显下降。1年生母树的插条生根率最高, 木质化枝和半木质化枝的生根率分别为42.3%和76.0%,4年生母树插条的生根率最低,分别为20.0%和67.8%。
|
|
对1年生母树6月20日扦插前未作任何处理的半木质化枝插条横切面进行观察,从外至内依次由周皮(含表皮残体)、皮层、次生维管组织3部分组成(图版Ⅰ-1)。周皮以内是皮层,该层细胞排列较疏松,细胞间有较大的空隙,细胞呈圆形,含有大量可见的结晶体。皮层内为维管组织系统,由韧皮部、维管形成层、木质部、维管射线和髓部组成。韧皮部外侧有数层厚壁细胞,排列成较为连续的环状。从茎的横切面上看,未发现潜伏根原基存在,说明在母株的正常生长过程中没有产生根原基。

|
图版Ⅰ Ar:不定根;Co:皮层; Le:皮孔;Pe:周皮; Ph:韧皮部;Pi:髓部;Ps:韧皮部厚壁组织; Rp:根原基; Vc:维管形成层; Ху:木质部。 Plate Ⅰ Ar: Adventitious root;Co: Cortex; Le: Lenticel; Pe: Periderm; Ph: Phloem; Pi: Pith; Ps: Phloem selerenchyraa; Rp: Root primordium; Vc: Vascular cambium; Xy: Xylem. 1. 1年生费约果半木质化枝横切面(40×); 2. 1年生费约果木质化枝横切面(40×); 3.2年生费约果半木质化枝横切面(40×); 4.2年生费约果木质化枝横切面(40×); 5.3年生费约果半木质化枝横切面(40×); 6.3年生费约果木质化枝横切面(40×); 7. 4年生费约果半木质化枝横切面(40×); 8.4年生费约果木质化枝横切面(40×); 9.根原基的形成(10×); 10.不定根伸出皮孔时的分层现象(10×); 11.不定根伸出皮孔(10×); 12.不定根。 1. Transverse section of soft branch of one-year-old mother tree (40×); 2. Transverse sectionof hard branch of one-year-old mothertree(40×); 3. Transverse section of soft branch of two-year-old mother tree (40×); 4. Transverse section of hard branch of two-year-old mother tree (40×); 5. Transverse section of soft branch of three-year-old mother tree (40×); 6. Transverse section of hard branch ofthree-year-old mother tree(40×); 7. Transverse sectionof soft branch of four-year-old mother tree(40×); 8. Transverse sectionof hardbranch of four-year-old mother tree (40×); 9. Development of root primordium (10×); 10. Delaminating phenomena of adventitiousroot coming out from lenticel (10×); 11. Adventitious root coming out from lenticel (10×); 12. Adventitious root. |
1) 不同树龄母树枝条解剖结构与插条生根的关系对6月20日采自1,2,3,4年生费约果母树的当年生半木质化枝茎横切面(图版Ⅰ-1,3,5,7)观察比较并测量(表 2),可以看出,不同树龄母树半木质化枝茎解剖结构以及表皮、皮层、韧皮部、木质部、髓半径的相对宽度和射线宽度等没有明显差异,形成层分生组织细胞层数都是4~5层。对采自1, 2, 3, 4年生费约果母树的当年生木质化枝茎横切面(图版Ⅰ-2,4,6,8)观察比较并测量,随着母树年龄的增大,木质化枝茎构造与半木质化枝构造没有发生明显的变化,形成层分生组织细胞层数也是4~5层。因此,从解剖学的角度看,随着费约果母树年龄的增大,其插条的生根能力不存在差异,费约果插条生根表现出的年龄效应是其他因素影响所致。
|
|
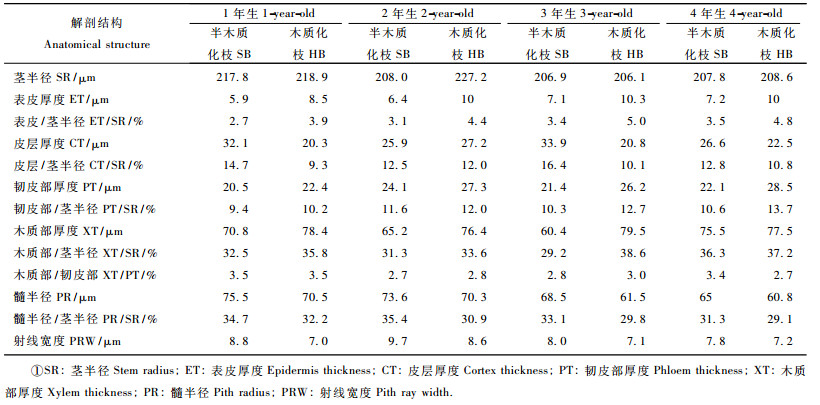
2) 同龄母树半木质化枝和木质化枝茎解剖结构与插条生根的关系对1年生费约果母树当年生半木质化枝和木质化枝茎结构进行比较发现,两者茎的组成部分没有变化,都是由周皮、皮层和维管系统三大部分组成,但各组成部分之间有着显著的差别(图版Ⅰ-1,2),从表 2可以看出:在整个切面上,表皮、木质部和韧皮部所占比例半木质化枝小于木质化枝,皮层所占比例半木质化枝大于木质化枝;髓在切面上占的比例半木质化枝大于木质化枝;射线宽度半木质化枝大于木质化枝;半木质化枝厚壁组织细胞稀疏且不连续,木质化枝厚壁组织细胞致密且连续。由于半木质化枝和木质化枝茎结构存在以上差异,可以推测半木质化枝插条较木质化枝插条生根能力强,因为木质化枝的表皮较半木质化枝厚,可能阻碍插条的不定根向外生长发育;木质化枝木质部所占比例较半木质化枝大,可塑性小,因而木质化枝分化能力较弱;木质化枝的厚壁组织细胞致密且连续,根原基突破厚壁组织困难,生根不易;半木质化枝的髓部大、射线宽,调节水分和营养物质的能力较强,有利于新根的形成。
2.3 费约果插条生根的解剖学观察6月20日扦插的1年生费约果半木质化枝插条在扦插后20天左右,形成层与髓射线细胞交叉处的薄壁细胞恢复分生能力,细胞质变浓,液泡缩小,细胞排列紧密,分裂形成一团细胞核较大、排列紧密并与周围细胞有明显区别的薄壁细胞团。这团薄壁细胞是从其先端(向着皮层的一端)不断分裂,并朝着根发端细胞发育方向伸展,分化出几个细胞核较大、细胞质较浓、染成红色的根原基位点细胞。根原基位点细胞及其附近形成层细胞染色较深,表现出强烈的分裂能力,逐渐形成体积较小、染色较深的一群分生细胞,构成分生组织团块(图版Ⅰ-9)。
分生组织细胞继续进行平周和垂周分裂,与根原基相连的、形成层周围的细胞分化较快,形成楔形的不定根原基。根原基沿着韧皮射线伸展方向发展,进入皮层(图版Ⅰ-10)。突破周皮时,不定根内部已出现明显的分层,维管系统分化完成,形成具有根冠、分生区及伸长区的根(图版Ⅰ-11)。随着不定根的不断生长发育,其颜色除根尖为白色外,其于部分逐渐变为褐色,且具有韧性,此时适宜移栽(图版Ⅰ-12)。
扦插20天后,插条基部形成愈伤组织,通过大量的观察,未见通过愈伤组织生出新根。
3 结论与讨论 3.1 扦插时期与生根的关系半木质化枝扦插较木质化枝扦插生根率高,不同扦插时期费约果半木质化枝插条的生根能力差异明显。在扦插环境条件得到保证的基础上,结合本试验研究结果,费约果在6月中旬左右采用半木质化枝扦插,适宜的扦插期约1个月。费约果随扦插时期的不同而引起的插条生根能力的差异,除与插条解剖结构有关外,可能还与插条内部激素水平和营养物质含量随生长周期而变化有密切关系。费约果扦插的最适时期,是以一定的生理状态为依据,而不是固定不变的日期,时期不同,生根情况也不同。
3.2 茎的解剖结构与不定根形成的关系费约果不定根的产生,必须有根原细胞的存在,在适宜的环境条件下,根原细胞分裂形成根原基,当根原基形成后,必须穿过韧皮部,突破皮层与韧皮部之间的较连续的环状厚壁组织到达皮层,最终伸出体外,形成不定根。费约果茎的解剖结构显示,次生韧皮部具有3~4层韧皮纤维细胞,细胞层数较多,且在次生结构中呈环状连续排列。王涛(1989)研究表明,在没有环状厚壁组织或厚壁组织不连续时,不定根的发生较容易,反之,若厚壁组织呈环状或多层则生根困难。刘卫东等(1997)在桃金娘科桉树(Eucalyptus spp.)上的研究表明,直杆桉(Eucalyptus maideni)插条生根困难可能与插条皮层内存在环状厚壁组织及幼枝中缺乏潜在根原基有关。易咏梅等(2000)的研究结果也证明,珙桐(Davidia involucrata)扦插生根困难与其茎的韧皮部存在一圈环状连续排列的厚壁细胞及插条内无原生根原基有关。王中英等(1986)指出,葡萄(Vitis vinifera)枝条中并无根原基,但其扦插容易生根,是其皮层薄、髓部大所决定的,植物枝条扦插由根原基、愈伤组织分化产生新根,枝条解剖结构也与扦插生根有密切关系。可见,费约果茎靠近韧皮部的皮层中的数层厚壁细胞是导致费约果扦插生根困难的主要原因之一。因此,费约果插条生根需时较长(约35天)可能与此有一定的关系。
3.3 插条年龄与生根的关系插条的生根能力是随着母树年龄的增长而降低的,在一般情况下母树年龄越大,植物插条生根就越困难,而母树年龄越小则生根越容易。插条年龄对生根的影响显著,一般以当年生枝的再生能力为最强,这是因为嫩枝插条内源生长素含量高、细胞分生能力旺盛,促进了不定根的形成。Tarrago(2005)等对大叶冬青(Ilex paraguariensis)嫩枝扦插的研究表明,来源于3年生母树插条的生根率显著高于20年生母树插条的生根率(P<0.05)。即使母树年龄差别很小,但其插条生根率可能存在较大差别,敖红等(2002)的研究结果表明,长白落叶松(Larix olgensis)2年生母树硬枝插条生根率(96.7%)显著高于其3年生母树的插条生根率(59.1%);沈海龙等(2005)指出,水曲柳(Fraxinus mandshurica)1年生实生苗插条平均生根率为83.0%,2年生为10.5%,3年生以上几乎不能生根。金庆等(2006)研究认为,随着母树年龄的增加,其插条生根率呈下降趋势。然而,王军辉(2006;2007)的研究表明,也有一些插条生根率受母树年龄影响较小或在特定年龄时生根较好的报道,这表明母树年龄效应在某些个别树种,尤其是难生根树种中并不是影响其插条生根的主导因子。本研究中,随着费约果母树年龄的增大,其插条的生根能力不存在差异,插条生根表现出的年龄效应是其他因素影响所致,与王军辉的研究结果一致。
3.4 费约果生根类型一般插条不定根根原基按形成的时间分为潜伏根原基和诱导根原基(刘勇等,1997)。潜伏根原始体多产生于维管系统内的韧皮组织、形成层或髓射线中,诱生根原始体是在扦插后才形成的,扦插前不存在根原基。许晓岗等(2006)在海棠果(Malus prunifolia)、王清民等(2006)在核桃(Juglans regia)上的扦插结果表明:两者为诱生根原基,初生射线与维管形成层交汇处细胞诱生根原基;刘云强等(2004)研究证实糠椴(Tilia mandshurica)的诱生根原基产生于维管形成层、愈伤组织;蒙椴(Tilia mongolica)的诱生根原基产生于髓射线与皮层交界处、维管形成层、愈伤组织。张晓平等(2004)在杂种鹅掌楸(Liriodendron chinense × L. tulipifera)上的研究表明,不定根原基起源于维管形成层区。郭素娟等(2004)研究表明,白皮松(Pinus bungeana)诱导根原始体,产生于维管形成层、韧皮组织与髓射线的交叉区域。本研究中,当年生费约果插条的茎中未发现潜伏的根原始体,且不定根是由扦插后形成的诱生根原基发育而来,因此,费约果插条的生根类型为诱导生根型。通过解剖观察,半木质化枝插条的根原基主要发生在维管形成层与髓射线交叉部位的薄壁细胞。解剖学观察发现,环状厚壁组织结构紧密,且多呈环状连续排列,这是费约果插条不易生根的原因之一。
敖红, 王昆, 冯玉龙. 2002. 长白落叶松插条的内源激素水平及其与扦插生根的关系[J]. 植物研究, 22(2): 190-195. |
郭素娟, 凌宏勤, 李凤兰. 2004. 白皮松扦插生根的生理生化基础的研究[J]. 北京林业大学学报, 26(3): 43-47. |
金庆, 秦峰, 储德裕, 等. 2006. 杂种马褂木扦插繁殖技术的研究[J]. 林业科学研究, 19(3): 370-375. |
李正理. 1987. 植物制片技术[M]. 北京: 科学出版社: 129-138.
|
刘勇, 肖德兴, 黄长干, 等. 1997. 板栗嫩枝扦插生根解剖学特征研究[J]. 园艺学报, 24(1): 8-12. |
刘卫东, 万朝琨, 饶龙兵, 等. 1997. 桉树扦插生根的解剖学研究[J]. 中南林学院学报, 17(4): 32-36. |
刘云强, 杨建民, 彭伟秀, 等. 2004. 两种椴树嫩枝扦插生根的解剖学研究[J]. 河北农业大学学报, 27(4): 33-37. |
沈海龙, 赵霞, 邢朝斌, 等. 2005. 水曲柳扦插繁殖影响因子的分析[J]. 东北林业大学学报, 33(3): 5-6. |
王丹, 刘仁道, 任少雄. 2007. 食用、观赏兼用果树新种类费约果引种的气候适应性分析[J]. 中国南方果树, 36(6): 39-41. |
王涛. 1989. 植物扦插繁殖技术[M]. 北京: 北京科学技术出版社: 57-58.
|
王军辉, 张建国, 张守攻, 等. 2006. 青海云杉硬枝扦插的激索、年龄和位置效应研究[J]. 西北农林科技大学学报:自然科学版, 34(7): 65-71. |
王军辉, 张建国, 张守攻, 等. 2007. 几种因素对青海云杉扦插繁殖生根的影响[J]. 南京林业大学学报:自然科学版, 31(1): 51-54. |
王清民, 彭伟秀, 吕保聚, 等. 2006. 核桃试管不定根的组织学研究[J]. 西北植物学报, 26(4): 719-724. |
王中英, 解思敏, 杨佩芳, 等. 1986. 葡萄的组织解剖构造研究[J]. 果树科学, 3(4): 26-28. |
许晓岗, 汤庚国, 童丽丽. 2006. 海棠果插条扦插生根过程解剖学观察[J]. 南京林业大学学报:自然科学版, 30(4): 77-80. |
易咏梅, 罗世家, 李鑫, 等. 2000. 珙桐茎的解剖构造及愈伤组织形成的研究[J]. 湖北民族学院学报, 18(3): 4-6. |
张晓平, 方炎明, 黄绍辉. 2004. 杂种鹅掌楸扦插生根过程中内源激素的变化[J]. 南京林业大学学报:自然科学版, 28(3): 79-82. |
Figueiredo S L B, Kersten E, Schuch M W. 1995. Effects of blanching and of indolbutyric acid (IBA) in the rooting of Feijoa sellowiana.Berg cuttings[J]. Scientia Agricola, 52(1): 167-171. DOI:10.1590/S0103-90161995000100029 |
Hardy P J, Micbapl B J. 1970. Volatile components of feijoa fruits[J]. Phytochemistry, 9: 1355-1357. DOI:10.1016/S0031-9422(00)85331-5 |
Kolesnik A A, Golubev V N, Gadzhieva A A. 1991. Lipids of the fruit of Feijoa sellowiana[J]. Chemistry of Natural Compounds, 27(4): 404-407. DOI:10.1007/BF00636556 |
Nakashima H. 2001. Biological activity of feijoa peel extracts.Occasional Papers[J]. Kagoshima University Research Center for the Pacific Islands, 34: 169-175. |
Tarrago J, Sansberro P, Filip R, et al. 2005. Effect of leaf retention and flavonoids on rooting of Ilex paraguariensis cuttings[J]. Scientia Horticulturae, 103: 479-488. DOI:10.1016/j.scienta.2004.07.004 |
Thorp G, Bieleskir R. 2002. Feijoas: Origins, cultivation and uses[M]. Auckland: David Bateman.
|