 2010, Vol. 46
2010, Vol. 46文章信息
- 张瑛春, 王军辉, 张守攻, 张建国, 孙晓梅, 朱景乐
- Zhang Yingchun, Wang Junhui, Zhang Shougong, Zhang Jianguo, Sun Xiaomei, Zhu Jingle
- Pilodyn和日本落叶松材性指标的关系
- Relationship between the Pilodyn Penetration and Wood Property of Larix kaempferi
- 林业科学, 2010, 46(7): 114-119.
- Scientia Silvae Sinicae, 2010, 46(7): 114-119.
-
文章历史
- 收稿日期:2008-12-19
- 修回日期:2009-06-21
-
作者相关文章



2. 甘肃小陇山林业实验局 天水 741022;
3. 中国林业科学研究院经济林研究开发中心 郑州 450003
2. Research Institute of Forestry of Xiaolongshan Tianshui 741022;
3. Non-Timber Forestry Research and Development Center, CAF Zhengzhou 450003
日本落叶松(Larix kaempferi)是我国北方及南方亚高山地区的重要针叶造林树种,具有早期速生、成林快、易于栽培、适应性广等特点(马常耕,1992); 且因其木材密度较其他针叶树种高,单位种植面积产木材量高,工业利用率高,是一种较好的造纸原料(周崟,2001; 王军辉等,2004)。木材密度是木材重要的物理性质指标,它直接影响木材的加工质量、制浆得率和浆料质量,并在很大程度上决定着木材其他的物理性质及力学性能。活立木密度测定的常规方法是在木材胸高处用生长锥取木芯,然后利用排水法测定木材基本密度,但这种方法不仅在一定程度上损坏树木,而且对于大批量样品的木材密度测定程序繁杂、耗时较长。Pilodyn是一种无损检测的仪器,可用于间接测定木材基本密度等材性指标。利用Pilodyn来测试活立木的木材密度不仅是一个迅速的、非破坏性、可替代传统取木芯进行测定的方法,同时还可用于木材密度的早期选择,加速林木育种进程(Wang et al., 2000; Cown,1978; Watt et al., 1996; Gough et al., 1984; 朱景乐等,2008a; 2008b)。但影响Pilodyn测定值的不仅仅是木材基本密度,还有林木生长量以及木材纤维特性等指标。本文通过Pilodyn对日本落叶松木材密度的测定研究,旨在探求Pilodyn测定值、木材基本密度、林木生长量和木材纤维特性间的关系,为日本落叶松木材利用、材性评价及遗传改良等工作提供间接木材密度评价指标,并探寻一种快速无损的测量木材密度及其他相关材性指标的方法。
1 材料与方法 1.1 试验地概况材料取自河南省西部伏牛山北坡的嵩县五马寺林场杨树岭林区日本落叶松种源试验林,33°34′—35°54′N,111°47′—112°15′E,海拔1 400 m,年均气温8.4 ℃, 7月份均温24.7 ℃, 1月份均温-5.5 ℃。无霜期178天,年降水量800~1 200 mm,空气相对湿度80%;土壤为棕壤,pH 6.0左右。
种源试验林定植于1993年,坡向为北坡,坡度20°,造林前试验地为栎类为主的落叶阔叶林采伐迹地。造林采用带状整地,株行距2 m×2 m。目前日本落叶松种源试验林内林木胸径在6~15 cm之间。
1.2 Pliodyn的工作原理本次试验选用的是6 J能量,2.5 mm直径探针型号的Pilodyn, 内径5 mm的生长锥。
Pilodyn的测定原理是以预先设定好的能量,将一个固定规格的探针射入到木材中,探针射入的深度就是检测的结果,这与木材密度及木材的纤维特征有密切负相关关系。木材密度大,则射入深度浅; 反之,射入深度深(Greaves et al., 1996; King et al., 1988; Micko et al., 1982; Moura et al., 1987; Talor,1981)。用所得到的Pilodyn探测数据和密度建立回归方程,采用建立的方程和其他活立木Pilodyn探测值预测对应活立木木材密度。
1.3 材性指标测定和测定方法在种源试验林内,按照随机取样原则选取日本落叶松79棵活立木。Pilodyn的取样方法是在胸高处分正南方向剥去活立木少量外部树皮, 用Pilodyn探测2次。如果2次探测值的误差在2 mm以内, 则取2次探测值的平均值作为南向Pilodyn的计算数据Ps; 如果2次探测值的误差大于2 mm, 则需要进行第3次南向Pilodyn探测,然后在3个数据中选择2个误差在2 mm以内, 且数据最接近的2个探测值的平均数作为Ps。用同样的方法测得北向的木材Pilodyn探测值Pn。
测定后在胸高处相同位置用内径5 mm的生长锥进行木芯取样,从树皮取到髓心,南北方向各取一个木芯进行室内材性指标的测定。室内材性指标包括南向整株木材基本密度(Ds)、北向整株木材基本密度(Dn)及其整株木材基本密度平均值(Da),早材管胞长度(FL1)、早材管胞宽度(FW1)、晚材管胞长度(FL2)和晚材管胞宽度(FW2),南向生长量(Gs)和北向生长量(Gn)。饱和含水量法测定木材密度,硝酸离析法测定管胞性状,每个样品均随机测定50个管胞值,并按其平均值进行计算。
1.4 数据统计分析方法将测试样本中不符合Pilodyn探测精度的数据剔除后,使用SPSS13.0软件对数据进行分析(张文彤等,2004)。
1) 比较南北方向林木生长量、木材密度、Pilodyn探测值间的差异情况, 分析木材基本密度、林木生长量、木材纤维特性及Pilodyn测定值间的相关关系。
2) 采用通径分析方法,解释各个因素对Pilodyn探测作用的理论依据, 并对通径分析进行检验,检验公式为W=-(n-d)lnQ,其近似服从自由度为d的χ2分布。n为样本数,d为待检验模型中减少的通径个数。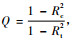
3) 随机采用49对数据通过回归分析方法建立木材密度等与Pilodyn测定值的回归方程, 并用剩余30对数据对该模型进行检验。模型为: Y=B+AX,预估精度检验公式为P= 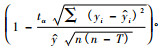
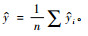
日本落叶松Pilodyn测定值、木材基本密度和林木生长量在南北方向上的遗传变异如表 1所示。由表 1可知,日本落叶松Ps(15.3)比Pn(14.6)高4.8%,Ds(0.351 0)略低Dn(0.353 0),Gs(4.780)比Gn(5.122)低6.7%。配对样本的t检验表明Pilodyn测定值、木材密度、林木生长量南北方向上都有显著差异。Pilodyn测定值、木材密度及林木生长量在各方向上变异系数比较接近,木材密度的变异系数显著小于其他2个指标的变异系数。
|
|
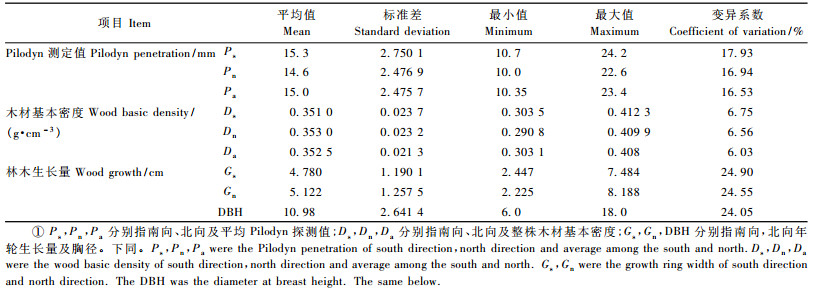
日本落叶松Pilodyn测定值、木材基本密度、木材纤维特性及生长量的相关关系如表 2所示。由表 2可知,Pilodyn测定值和相同方向上木材密度存在极显著的负相关(-0.454~-0.587),Pilodyn和相同方向上的林木生长量存在极显著的负相关(-0.394~-0.459),Pilodyn和晚材的纤维特性存在极显著的负相关(-0.329~-0.447),而与早材的纤维特性没有显著的相关性(-0.015~-0.137)。
|
|
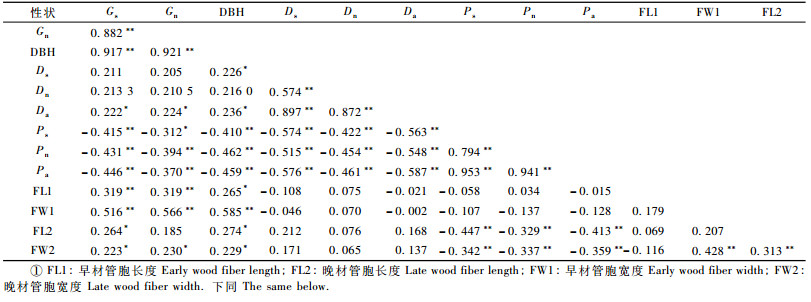
木材密度与相同方向上林木生长量没有显著相关(0.210 5~0.211 0),木材密度与纤维特性无显著相关(-0.021~0.212)。
纤维特性和林木生长量上存在显著正相关(0.185~0.585)。纤维性状间仅晚材管胞宽度和早材管胞宽度、晚材管胞长存在极显著的相关关系(0.313~0.428),其他纤维特性间均没有显著的相关关系(-0.116~0.179)。
整株木材基本密度和林木的年轮南北向生长量存在显著的相关关系(0.222~0.224),胸径和南北方向的木材密度的相关关系在临界值0.221(df=77,α=0.05)附近(0.216~0.236),南北方向上木材密度和对应的年轮生长量的相关关系没有达到显著水平(0.210 5~0.211),不过和临界值相差不大,所以尚需进一步研究才能确定其相关关系。
2.3 通径分析为了研究Pilodyn测定值与林木生长和材性的关系,以及其他材性指标如何作用于Pilodyn测定值,需要用各个因子对Pilodyn测定值做通径分析,求出各个因子的贡献率。按照生物学意义构建各个因子对Pilodyn测定值的通径关系如图 1A所示,但因通径链太多,且有些通径链没有统计意义,这样不仅增加计算的难度,同时对于Pilodyn的工作原理的揭示也存在一定的误差。因此将其中不显著的通径链删除后,得到的简化模型如图 1B所示。按照前文提供的方法对图 1B做检验,图 1B对图 1A的Q值为85.22%,W值为10.869,小于置信空间为0.05、自由度为12的卡方值,认为图 1B能够提供图 1A中85.22%的信息,因此简化是可行的。

|
图 1 各指标对Pilodyn测定值的通径图 Figure 1 The path graphics of Pilodyn penetration with each wood index |
用缩减后的模型来对各个因子对Pilodyn测定值的作用结果做深入研究。各个因子对Pilodyn测定值的直接和间接作用如表 3所示。由表 3可知,各个因素对于Pilodyn测定值的总作用与该因素与Pilodyn测定值相关系数的差值即为删除掉没有显著统计学意义的通径链间的总作用。南向木材基本密度对Pilodyn测定值直接通径系数是-0.462,与相关系数(-0.574)差别不是很大(-0.112),所以认为木材密度对于Pilodyn测定值作用的解释基本合理。早材管胞长和早材管胞宽对于Pilodyn测定值没有直接的作用关系,因此模型总作用即为其间接作用,与相关系数几乎相同, 所以认为早材管胞长、早材管胞宽对于Pilodyn没有显著的作用。
|
|
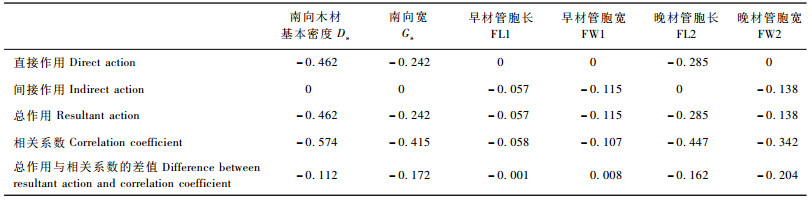
南向木材基本密度对Pilodyn测定值贡献率最大(46.71%),晚材管胞长和南向生长量贡献率次之,依次为28.82%和24.47%,其余指标对Pilodyn测定值均没有直接作用。因此当排除木材基本密度及南向生长量对Pilodyn测定值影响后,使用Pilodyn测定值进行晚材管胞长的预测也是有可能的。
2.4 木材密度的预测及方程精度检验因为木材密度对于Pilodyn测定值的贡献最大,所以使用Pilodyn测定值预测木材密度是可行的。使用49套数据作为拟和样本,建立Pilodyn测定值和木材密度线性回归关系如图 2所示。由图 2可知,平均值y= -0.005 5x + 0.437 4 (R2=0.393 1),北向: y=-0.004 1x+0.412 5(R2=0.209), 南向: y=-0.006 1x+0.447 5(R2=0.425 3),南向的决定系数高于北向的决定系数,因此在南向取得的Pilodyn数据比在北向取得的Pilodyn数据更能反映木材密度的变化。使用剩余的30套数据作为检验样本,对拟合的3个方程进行预估精度计算,得到的预估精度分别为南向98.05%,北向97.80%,平均98.31%,均超过95%,因此该模型用来预测日本落叶松得木材基本密度是可行和可靠的。

|
图 2 Pilodyn测定值和对应方向上木材密度的线性回归方程 Figure 2 The linear regression equation between the Pilodyn penetration and wood basic density in the relevant direction |
日本落叶松Ps(15.3)高于Pn(14.6)4.8%,Ds(0.351 0)略低于Dn(0.353 0),Gs(4.780)比Gn(5.122)低6.7%,配对样本t检验结果表明Pilodyn测定值、木材密度、生长量在南北方向上都有显著差异。该结论与Muneri等(2001)通过对蓝桉(Eucalyptus globulus)、亮果桉(E. nitens)的研究,不同方向上木材的纤维宽度和长度没有显著差异,树木南北方向的木材密度没有显著差异不同,而与王慧梅等(2004)在红松(Pinus koraiensis)上研究得到南北方向上年轮生长量有显著差异的结果一致。
3.2 日本落叶松各性状间的相关关系Pilodyn测定值和相同方向上木材密度存在极显著的负相关(-0.454~-0.587),Cown(1978)研究表明辐射松(Pinus radiata)Pilodyn探测值与其木材密度呈显著负相关(-0.96), 白云杉(Picea glauca)Pilodyn测定值和密度也呈显著负相关(-0.83)(Micko et al., 1982)。加拿大Hall在18个不同林龄的白云杉、黑云杉(Picea mariana)和北美落叶松(Larix laricina)中大批量试用,并与传统排水法所测的结果相比较以检验其可用性和范围,看到Pilodyn的读数与直测木材密度间相关系数因树种变动在-0.451~-0.564之间(马常耕, 1998)。Pilodyn测定值和木材密度间显著的相关关系为Pilodyn测定值快速预测日本落叶松木材密度提供了依据。
Pilodyn测定值和木材纤维特性的关系比较复杂,Pilodyn测定值和早材的纤维特性无显著的相关,Pilodyn测定值和晚材的纤维特性存在显著的相关关系,因为晚材的质地比较紧密,早材的质地疏松,他们对Pilodyn的阻力不同造成了他们之间紧密关系的不同。
Pilodyn测定值和相同方向上的林木生长量存在极显著负相关(-0.394,-0.459),纤维特性和林木生长量上存在显著正相关(0.185~0.585)。与孙晓梅等(2005)在日本落叶松上得到的纤维长度与木材生长量存在显著的相关关系结果相同。
木材密度与纤维特性的相关关系不显著, 这与孙晓梅等(2005)在日本落叶松上得到木材密度和木材的纤维特性没有显著相关关系,两者是相互独立的,王军辉等(2001)在桤木(Alnus cremastogyne)上得到木材密度和早材纤维特性没有显著的相关关系的结果一致。
3.3 通径分析根据生物学意义建立各个指标对于Pilodyn测定值的通径图,并对不显著通径链进行缩减,经检验得到的模型和原模型没有显著差异。在新模型中得到木材的基本密度、年轮生长量以及晚材纤维长度均能直接作用于Pilodyn测定值,早材纤维特性是通过生长量间接作用于Pilodyn测定值,但纤维特性却和木材的基本密度没有显著的作用。通径分析的结果验证了日本落叶松木材纤维特性和木材密度间相互独立,木材年轮生长量和木材基本密度相互独立,本研究结果和Raymond等(1998)的研究表明树木的密度和纤维特性与直径相关关系小,研究这方面无意义。Moura等(1987)研究表明木材密度与胸径大小只有弱的相关关系; King等(1988)指出,木材密度和直径有强的负相关关系。Wang等(2000)对小干松(Pinus contorta var. latifolia)研究中生长轮宽度与相应木材密度呈负相关关系; 王慧梅等(2001)对红松的研究中得到,年轮宽度与木材密度存在显著的相关关系(r=-0.67), 年轮越窄,木材密度越大; 反之,则木材密度越小。
木材基本密度对Pilodyn测定值的直接通径系数是-0.462,贡献率最大(46.71%)。林木生长量对Pilodyn测定值的直接通径系数为-0.242,,贡献率为24.47%,晚材管胞长直接通径系数为-0.285,贡献率为28.82%。
纤维特性指标与木材密度、生长量与木材密度间均没有通径链,表明木材密度与纤维特性、木材密度与生长量间是相互独立的,这为日本落叶松生长和材性的联合改良提供了依据。与前人已经证明了的杨树材质性状指标和生长性状间是相互独立的,可进行联合选择的结果一致(王克胜等,1995; 李善文等,2005)。
3.4 对2个方向上木材基本密度和2个方向木材基本密度平均值进行方程拟和,并进行检验因为木材密度对于Pilodyn测定值的贡献最大,所以使用Pilodyn测定值预测木材密度是可行的。使用拟合样本得到的方程中南向的决定系数(0.425 3)高于北向的决定系数(0.209),因此在南向进行Pilodyn测量取得的数据比在北向取得的Pilodyn数据更能反映木材密度的变化。使用检验样本对3个方程进行检验,得到的预估精度分别是南向98.05%,北向97.80%,平均98.31%,均超过95%,因此该模型用来预测日本落叶松木材基本密度是可行和可靠的。南向Pilodyn测定值与木材密度和纤维特性的相关关系均高于北向的,因此在南向进行Pilodyn测定能取得较好的效果。
郭志刚. 1999. 社会统计分析方法[M]. 北京: 中国人民大学出版社: 145-174.
|
李善文, 姜岳忠, 王桂岩, 等. 2005. 黑杨派无性系生长与材性联合选择[J]. 林业科学, 41(2): 53-58. DOI:10.11707/j.1001-7488.20050209 |
马常耕. 1992. 落叶松种和种源选择[M]. 北京: 中国农业大学出版社.
|
马常耕. 1998. 国外制浆材性状的测定技术[J]. 世界林业研究, 5: 38-43. |
孙晓梅, 张守攻, 李时元, 等. 2005. 日本落叶松自由授粉家系纸浆材材性遗传变异的研究[J]. 林业科学, 41(4): 48-54. DOI:10.11707/j.1001-7488.20050409 |
王克胜, 韩一凡, 任建中, 等. 1995. 群众杨改良无性系材性的遗传及性状相关的研究[J]. 林业科学, 31(1): 44-49. |
王慧梅, 夏德安, 王文杰. 2004. 红松种源材质性状研究[J]. 植物研究, 24(4): 495-498. |
王军辉, 陆熙娴, 张守攻. 2004. 落叶松制浆造纸适应性及开发利用的前景[J]. 中国造纸, 23(6): 47-52. |
王军辉, 顾万春, 夏良放, 等. 2001. 桤木种源(群体)/家系材性性状的遗传变异[J]. 林业科学研究, 14(4): 362-368. |
张文彤, 董伟., 等. 2004. SPSS统计分析高级教程[M]. 北京: 高等教育出版社: 137-148.
|
周崟. 2001. 中国落叶松属木材[M]. 北京: 中国林业出版社: 166-173.
|
朱景乐, 王军辉, 张守攻, 等. 2008. Pilodyn在林木遗传改良中的研究进展[J]. 浙江林学院学报, 25(5): 661-665. |
朱景乐, 王军辉, 张守攻, 等. 2008. 毛白杨材性指标预测与选择[J]. 林业科学, 44(7): 23-28. DOI:10.11707/j.1001-7488.20080705 |
Cown D J. 1978. Comparison of the Pilodyn and torsiometer methods for the rapid assessment of wood density in living trees[J]. N Z J For Sci, 8: 384-391. |
Gough G, Barbes R D. 1984. A comparison of three methods of wood density assessment in a Pinus elliottii progeny test[J]. South African Forestry, 128: 22-25. DOI:10.1080/00382167.1984.9628921 |
Greaves B L, Borrslho N M G, Raymond C A, et al. 1996. Use of a Pilodyn for the indirect selection of basic density in Eucalyptus nitens[J]. Canadian Journal of Forest Research, 26: 1643-1650. DOI:10.1139/x26-185 |
King J N, Yeh F C, Heaman J C, et al. 1988. Selection of wood density and diamter in controlled crosses of coastal Douglas-fir[J]. Silvae Genetica, 37: 152-157. |
Micko M M, Wang E I C, Taylor F W, et al. 1982. Determination of wood density in standing white spruce using Pilodyn tester[J]. The Forestry Chronicle, 58(4): 178-180. DOI:10.5558/tfc58178-4 |
Moura V P G, Barnes R D, Birks J S. 1987. A comparison of three methods of assessing wood density inprovenances of Eucalyptus camaldulensis Dehn.and other Eucalyptus spp. in Brazil[J]. Australian Forest Research, 17: 83-90. |
Muneri A, Raymond C A. 2001. Nondestructive sampling of Eucalyptus globulus and E.nitens for wood properties. Ⅱ. Fibre length and coarseness wood[J]. Wood Science and Technology, 35(1-2): 41-56. DOI:10.1007/s002260100088 |
Raymond C A, Macdonald A C. 1998. Where to shoot your Pilodyn: within tree variation in basic density in plantation Eucalyptus globulus and E.nitens in Tasmania[J]. New Forest, 15(3): 205-221. DOI:10.1023/A:1006544918632 |
Talor F W. 1981. Rapid determination of southern pine specific gravity with a Pilodyn tester[J]. Forest Science, 27: 59-61. |
Wang T L, Aitken S N. 2000. Selection for improved growth and wood density in lodgepole pine: effects on radial patterns of wood variation[J]. Wood Fiber Sci, 32(4): 391-403. |
Watt M S, Garnett B T, Walker J C F. 1996. The use of the Pilodyn for assessing outerwood density in New Zealand radiata pine[J]. Forest Products Journal, 46: 11-12, 101-106. |