 2010, Vol. 46
2010, Vol. 46文章信息
- 褚延广, 苏晓华, 黄秦军, 张香华
- Chu Yanguang, Su Xiaohua, Huang Qinjun, Zhang Xianghua
- 欧洲黑杨基因资源光合生理特征与生长的关系
- Relationships between Photosynthetic Characteristics and Growth Traits in Gene Resources of Populus nigra
- 林业科学, 2010, 46(7): 77-83.
- Scientia Silvae Sinicae, 2010, 46(7): 77-83.
-
文章历史
- 收稿日期:2009-04-13
- 修回日期:2010-04-22
-
作者相关文章



光合作用是影响植物生长的关键因素之一,其主要表现在植物体干物质积累过程对光合作用的依赖。因此,光能利用效率的高低直接影响植物生物量的大小(侯爱菊等,2005)。由于培育光能利用率高的速生杨树(Populus)新品种是杨树育种的重要目标,光合生理特征也就成为对杨树新品种进行评价的重要指标,对研究如何提高杨树品种对光能资源的利用效率也具有重要的理论意义和经济意义。
对净光合速率(Pn)等植物气体交换相关参数的测定,可为评价植物光合生理特征提供依据;此外,叶绿素荧光检测技术也是分析植物光合能力的重要手段,可为研究光系统Ⅱ提供丰富的信息(黄焱等,2008)。以往对杨树光合生理的研究仅限于对特定品种的评价,对杨树基因资源的气体交换与叶绿素荧光特征的研究则少见报道。欧洲黑杨(Populus nigra)属杨属(Populus)、黑杨派(Aigeiros)(Eckenwalder,1996),广泛分布于欧洲、非洲北部、亚洲中部和西部(Vanden,2003),我国仅新疆北部有少量分布。欧洲黑杨也是杨树杂交育种中重要的基因供体。目前已对欧洲黑杨的木材材性、生长、叶面积和稳定碳同位素组成等特征进行了资源评价研究(丁明明,2006;丁明明等,2006),但对其光合生理特征的了解还很有限。本文研究引自国外及国内不同地区欧洲黑杨无性系(基因型)与光合生理相关的气体交换和叶绿素荧光特征的遗传变异情况及其与生长性状的关系,初步评价欧洲黑杨基因资源的光合特性,以期为杨树高光效新品种选育提供优良的亲本材料。
1 材料与方法 1.1 材料与试验设计本研究所用欧洲黑杨共108个无性系(基因型),分别为2000年起引自欧洲黑杨分布区的14个欧洲国家(南欧:罗马尼亚、塞尔维亚、西班牙、意大利、克罗地亚、土耳其;东欧:俄罗斯;西欧:英国、荷兰、比利时;中欧:捷克、斯洛伐克、德国、匈牙利)及我国新疆西北部。各参试无性系产地地理及气候资料见表 1。
|
|
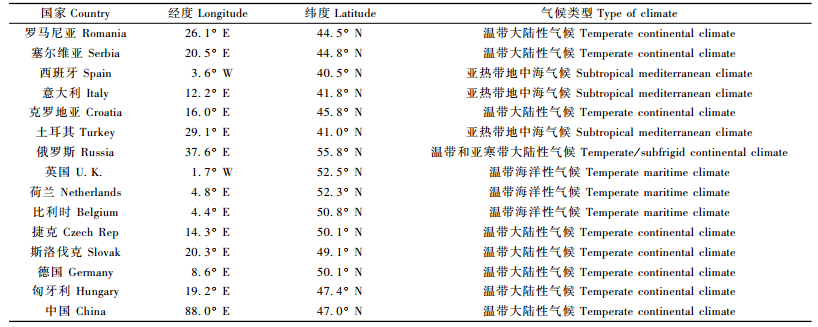
2007年春季,取各参试无性系1年生苗,将茎剪成15 cm长的插穗,扦插于北京房山区组村苗圃(39°34′ N,116°05′ E)。采用完全随机区组设计,4次重复,6株小区,行间距90 cm,株间距60 cm。试验地采用常规方法进行管理,定期清理杂草、人工去除侧枝、定期浇水灌溉。试验地内不进行施肥操作,不使用农药。
1.2 气体交换参数测定于2007年7月对1年生欧洲黑杨扦插苗生长旺盛时进行光合特性的测定。测定工作在7月21,22,23日连续的3天内完成,测定时间为每天上午8:30—11:30,测定期间试验地所在地天气晴朗,气温和湿度稳定。试验采用美国LI-COR公司Li-6400型便携式光合仪进行,并使用人工光源(光强2 000 μmol·m-2s-1)和CO2注入系统。钢瓶设定CO2注入浓度为400 μmol·mol-1,叶室相对湿度为70%~80%,测定叶片温度为25~32 ℃。每株待测植株上取3片功能叶进行检测,每个无性系取3株树进行测定。所记录瞬时气体交换参数包括净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)等。
1.3 叶绿素荧光参数及生长测定于2007年7月对1年生欧洲黑杨扦插苗生长旺盛时进行叶绿素荧光特性检测,检测在气体交换参数测定后的7月24,25,26日3天内完成,测定时间为每天上午8:30—11:30。采用德国Walz公司PAM-2100型便携式荧光仪进行测定。检测在上午8:30—11:30期间、天气晴朗、气温、湿度适中的条件下进行,饱和脉冲光强为4 500 μmol·m-2s-1,光化光强为800 μmol·m-2s-1。所测定参数包括PSⅡ电子传递速率(ETR)、PSⅡ实际量子产量(Yield)和PSⅡ最大光化学效率(Fv/Fm)(Maxwell et al., 2000)。Yield的计算公式为:Yield=(Fm'-Ft)/Fm',其中Fm'为光适应下最大荧光,Ft为任一给定时间的实际荧光;ETR计算公式为:ETR=Yield×PAR×0.5×0.84,其中PAR为光合有效辐射强度;Fv/Fm的计算公式为:Fv/Fm=(Fm-Fo)/Fm,其中Fo,Fm和Fv分别为暗适应下初始荧光、最大荧光和可变荧光;各参试无性系植株取第7片功能叶测定,每个无性系取3株树进行测定,测定前采用叶片夹进行20 min遮光处理,以保证叶片完全进入暗适应状态。对每株已进行气体交换和叶绿素荧光参数测定的欧洲黑杨测量其树高和胸径等生长性状。
1.4 数据分析采用SPSS13.0软件包(SPSS Inc.,USA)进行统计分析。采用单因素方差分析计算各参数指标在试验群体内的差异程度;各参数广义遗传力(broad sense heritability,H2)分析方法见(Singh et al., 1993),即为基因型变量(σg2)与表现型变量(σg2+σe2)的比值:H2=σg2/(σg2+σe2)。变异系数采用公式C=S/X计算;式中S为标准差,X为某一指标的总体平均值。采用相关分析分析各参数指标及无性系产地经度、纬度间的相关关系。
2 结果与分析 2.1 欧洲黑杨气体交换、叶绿素荧光与生长特征及其变异以栽植于北京的108个欧洲黑杨1年生植株为材料,测定其气体交换、叶绿素荧光及树高、胸径生长特征值。由表 2可见:除Fv/Fm值外,无性系间其他各参数测量值均具有较大的变幅。方差分析结果表明,无性系间气体交换参数Pn,Ci,Gs和Tr,荧光参数ETR和Yield值的差异均极显著水平(P<0.001),而Fv/Fm值间的差异不显著(P=0.172)。
|
|
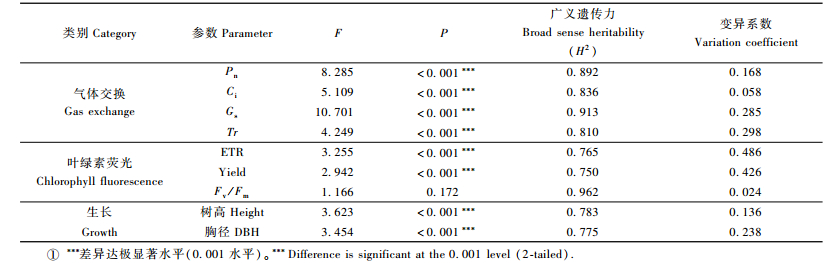
表型性状的遗传力特征可为利用性状变异进行优良基因型的选择提供参考。分析结果显示:各参数均具有较高的广义遗传力,其中气体交换参数的广义遗传力水平总体高于叶绿素荧光参数;Fv/Fm值具有最高的遗传力(H2=0.962),说明该性状受遗传控制最强,受环境影响最小。各参数的变异系数差别较大,除Ci和Fv/Fm较小外(分别为0.058和0.024),其他参数均具有较大的变异系数(0.136~0.748),说明欧洲黑杨基因资源气体交换、叶绿素荧光、生长性状具有较大的遗传潜力。
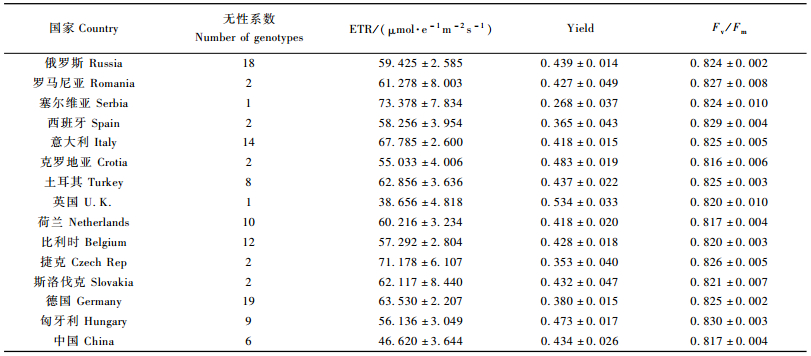
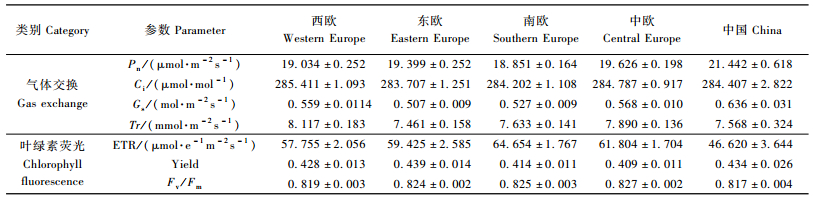
2.2 不同国家、不同地区间欧洲黑杨气体交换及叶绿素荧光特征的变异分析结果发现:不同国家无性系气体交换主要参数Pn,Ci,Gs和Tr的测定值范围分别为(17.739±0.647)~(21.442±0.618)μmol·m-2s-1,(275.333±7.259)~(293.611±5.365)μmol·mol-1,(0.486±0.034)~(0.651±0.016)mol·m-2s-1和(6.867±0.563)~(8.733±0.715)mmol·m-2s-1(表 3);叶绿素荧光参数ETR,Yield和Fv/Fm测定值范围分别为(38.656±4.818)~(67.785±2.600)μmol·e-1m-2s-1,(0.268±0.037)~(0.534±0.033)和(0.816±0.006)~(0.830±0.003)(表 4)。说明不同基因型在这些性状上存在广泛的变异。从不同国家来看:来自我国和英国的欧洲黑杨基因资源具有较高的Pn值,而引自西班牙的欧洲黑杨的Pn值最低;引自英国的欧洲黑杨还具有最高的Ci和Yield值,而引自匈牙利的欧洲黑杨的Fv/Fm值最高。
|
|
|
|
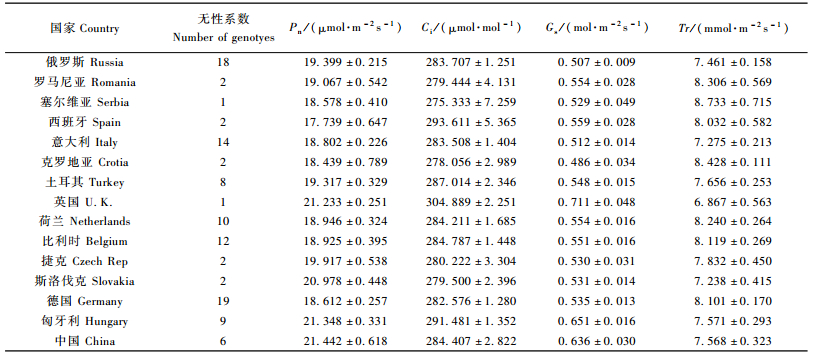
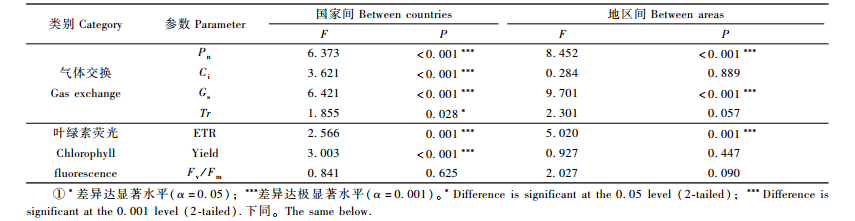
从不同分布区来看:来自我国的欧洲黑杨基因资源具有较高的Pn和Gs值,引自中欧地区的欧洲黑杨则具有较高的Fv/Fm值(表 5)。方差分析(表 6)结果表明:各国家间欧洲黑杨基因资源的Pn,Ci,Gs,ETR和Yield等气体交换与叶绿素荧光参数差异均达极显著水平;各分布区域间欧洲黑杨Pn,Gs和ETR等参数的差异也均达极显著水平,说明欧洲黑杨基因资源在其分布区内气体交换与叶绿素荧光相关性状变异程度较大,可为进一步的杨树高光效育种研究提供丰富的资源基础。
|
|
|
|
相关分析结果见表 7。分析结果显示:欧洲黑杨净光合速率(Pn)与气孔导度(Gs)呈极显著正相关,与PSⅡ电子传递速率(ETR)呈极显著负相关,说明气孔导度较高的欧洲黑杨基因型具有较高的净光合速率,而其PSⅡ电子传递速率则可能较低。Fv/Fm值仅与ETR呈显著正相关,而与净光合速率等其他参数无相关关系,表明从欧洲黑杨自身,PSⅡ电子传递速率可能是决定其最大光化学效率的主要因子,电子传递速率较高的基因型具有较高的PSⅡ最大光化学效率。
|
|

气体交换参数Pn,Ci,Gs分别与树高呈显著负线性相关,叶绿素荧光参数ETR与树高呈显著正线性相关,除树高外的其他参数与胸径无明显的相关性(表 7,图 1)。由于高生长与树木木材产量紧密相关,因此Pn,Ci,Gs和ETR在对欧洲黑杨光合利用效率的评价中具有潜在的应用价值。结合表 3, 4结果可知:起源于塞尔维亚的欧洲黑杨具有较低的Pn,Ci,Gs值和较高的ETR值,推测该国的欧洲黑杨可能具有较高的光合利用效率。从地区的角度来看(表 5),起源于南欧和东欧的欧洲黑杨可能具有高的光合利用效率。欧洲黑杨气体交换、叶绿素荧光、生长特征参数与各无性系的原产地地理位置(经度和纬度)无明显的相关关系,提示经纬度不是影响欧洲黑杨光合能力的关键因素。

|
图 1 欧洲黑杨气体交换、叶绿素荧光参数与树高的相关性 Figure 1 The relationships among gas exchange, chlorophyll fluorescence and height in P. nigra |
目前,杨树光合生理特征的研究多集中在对品种的生长与抗逆性等性状的评价方面。如付士磊等(2006)研究干旱胁迫下小青杨(P. pseudo-simonii)的光合生理变化;万雪琴等(2008)探讨重金属镉(Cd)处理对3个欧美杨无性系叶绿素荧光参数的影响;吴瑞云等(2008)研究空气干旱对美洲黑杨“中潜6”(P. deltoides Cl. ‘Zhongqian6’)夏季和秋季光合生理的影响。以杨树基因资源为对象进行气体交换与叶绿素荧光特征评价的研究还很少,如周洪华等(2008)曾报道塔里木河下游胡杨(P. euphratica)天然林的光合特性,而将气体交换、叶绿素荧光特征与生长性状结合分析的研究还未见报道。
植物光合效率的高低,直接影响个体有机物积累的多少。以提高产量为目标的高光效育种研究在作物中已有许多探索(侯爱菊等,2005;陈悦等,2007;Long et al., 2006),林木中相关的理论与实践研究还十分缺乏。以为杨树高光效育种提供基础为目标,本文对引自欧洲及我国新疆的欧洲黑杨基因资源的气体交换、叶绿素荧光参数、生长特征进行测定与分析。由于所有参试无性系(基因型)在同一试验点栽植,在相同生长条件下进行测试,因此所测表型值的差异主要来源于遗传因素。试验所得气体交换与叶绿素荧光参数虽为瞬时值,但仍可反映基因资源内光合能力的基本情况。
本文的结果表明:起源于塞尔维亚的欧洲黑杨可能具有较高的光合利用效率,及其光合作用对产量的贡献较大;起源于塞尔维亚及其他南欧、东欧的欧洲黑杨可在进一步的杨树高光效育种研究中作为优良基因型选择的重点。与之前的报道相比(周永学等,2004),欧洲黑杨气体交换、叶绿素荧光特征与生长性状具有相似水平的广义遗传力,但本文结果中其变异系数则普遍较高。气孔导度Gs与净光合速率Pn,Ci呈极显著正相关,ETR与Pn呈极显著负相关,暗示Gs和ETR可能是影响欧洲黑杨光合能力的重要限制因素。林木的单株树高不仅与单位面积净光合速率有关,还与总同化面积即叶面积有关,欧洲黑杨无性系Pn与树高呈显著负相关,而叶面积与树高呈显著正相关(丁明明,2006),说明较大同化面积可是Pn较低的欧洲黑杨基因资源具有较大高生长的重要因素。ETR的高低直接影响ATP和NADPH等同化力的产生,但表观电子传递速率还受环式光合电子传递、抗坏血酸电子传递等因素有关,因此欧洲黑杨ETR与Pn负相关的原因还需进一步研究。此外,Fv/Fm值在无性系间的差异不显著(表 2,表 6),且与多数参数无相关关系,说明其在欧洲黑杨群体内的变异较小、遗传稳定性高。PSⅡ的最大量子产量Fv/Fm反映植物光系统Ⅱ(PSⅡ)的光化学效率,因此成为评价PSⅡ有效产率或最大光化学效率的重要参数(Maxwell et al., 2000;Schreiber et al., 2004)。无性系间Fv/Fm值细微的差别对欧洲黑杨光能利用效率的影响如何,还需要进一步进行研究。
本文的结果表明:气体交换与叶绿素荧光参数在欧洲黑杨中是高度受遗传控制的,因此在杨树高光效新品种培育研究中具有现实的意义。欧洲黑杨基因资源气体交换与叶绿素荧光具有丰富的变异,且部分参数与生长性状显著相关,可为进一步优良亲本材料的选择和杨树高光效新品种的培育提供理论基础与基因资源。
陈悦, 袁隆平, 王学华, 等. 2007. Relationship between grain yield and leaf photosynthetic rate in super hybrid rice[J]. 植物生理与分子生物学学报: 235-243. |
丁明明, 苏晓华, 黄秦军. 2006. 欧洲黑杨基因资源稳定碳同位素组成特征[J]. 林业科学研究, 19(3): 272-276. |
丁明明. 2006. 欧洲黑杨育种基因资源评价研究. 中国林业科学研究院博士学位论文. http://d.wanfangdata.com.cn/Thesis/Y962173
|
付士磊, 周永斌, 何兴元, 等. 2006. 干旱胁迫对杨树光合生理指标的影响[J]. 应用生态学报, 17(11): 2016-2019. DOI:10.3321/j.issn:1001-9332.2006.11.004 |
侯爱菊, 徐德昌. 2005. 植物高光效基因工程育种[J]. 中国生物工程杂志, 25(9): 19-23. |
黄焱, 季孔庶, 汤庚国. 2008. 珍珠黄杨不同扦插继代的荧光特性[J]. 东北林业大学学报, 36(10): 9-11, 15. DOI:10.3969/j.issn.1000-5382.2008.10.003 |
万雪琴, 张帆, 夏新莉, 等. 2008. 镉处理对杨树光合作用及叶绿素荧光参数的影响[J]. 林业科学, 44(6): 73-78. DOI:10.11707/j.1001-7488.20080612 |
吴瑞云, 郑红波, 王少元, 等. 2008. 空气干旱对美洲黑杨夏秋两季光合生理的影响[J]. 现代农业科学, 15(6): 30-32. |
周洪华, 陈亚宁, 李卫红, 等. 2008. 塔里木河下游胡杨气体交换特性及其环境解释[J]. 中国沙漠, 28(4): 665-672. |
周永学, 苏晓华, 樊军峰, 等. 2004. 引种欧洲黑杨无性系苗期生长测定与选择[J]. 西北农林科技大学学报:自然科学版, 32(10): 102-106. |
Eckenwalder J E. 1996.Systematic and evolution of Populus//Stettler R F, Bradshaw H D Jr, Heilman P E, et al. Biology of Populus and its implications for management and conservation. NRC Research Press, National Council of Canada, Ottawa, 7-32.
|
Long S P, Zhu X G, Naidu S L, et al. 2006. Can improvement in photosynthesis increase crop yields?[J]. Plant Cell Environ: 312-330. |
Maxwell K, Johnson G N. 2000. Chlorophyll fluorescence―a practical guide[J]. J Exp Bot, 51(345): 659-668. DOI:10.1093/jexbot/51.345.659 |
Schreiber U. 2004. Pulse Amplitude (PAM) fluorometry and saturation pulse method//Papageorgiou G, Govind Jee. Chlorophyll fluorescence: A signature of photosynthesis. Kluwer Academic Publishers, Dordrecht, The Netherlands, 279-319.
|
Singh M, Ceccarelli S, Hamblin J. 1993. Estimation of heritability from varietal trials data[J]. Theor Appl Genet, 86(4): 437-441. DOI:10.1007/BF00838558 |
Vanden B A. 2003. EUFORGEN technical guidelines for genetic conservation and use for european black poplar (Populus nigra L.)[J]. International Plant Genetic Resources Institute, Rome: 1-6. |