 2010, Vol. 46
2010, Vol. 46文章信息
- 李修鹏, 俞慈英, 吴月燕, 洪中跃, 孙晶, 陈叶平, 缪玲霞
- Li Xiupeng, Yu Ciying, Wu Yueyan, Hong Zhongyue, Sun Jing, Chen Yeping, Miao Lingxia
- 普陀鹅耳枥濒危的生物学原因及基因资源保存措施
- The Biological Reason for Endangerment of Carpinus putoensis and Measures for Gene Conservation
- 林业科学, 2010, 46(7): 69-76.
- Scientia Silvae Sinicae, 2010, 46(7): 69-76.
-
文章历史
- 收稿日期:2009-08-10
- 修回日期:2009-11-25
-
作者相关文章


2. 浙江省舟山市林业科学研究所 舟山 316000;
3. 浙江万里学院 宁波 315100
2. Zhoushan Forestry Institute, Zhejiang Province Zhoushan 316000;
3. Zhejiang Wanli University Ningbo 315100
物种的成功保育在于保育物种的基因库,即保育其最大的遗传多样性,以提供物种应对环境变迁的持续进化潜力。植物野生种群的遗传结构与其生态生物学特性有密切关系(Lande,1988;李忠超等,2006), 研究植物的生物学、生态学特性与繁殖机制,不但可以了解物种的濒危机制,为就地保育如何保护种群遗传基础和不同生态位的生态适应性提供依据,还将为迁地保育和野外重建的有效性提供资料(李忠超等,2006)。
普陀鹅耳枥(Carpinus putoensis)为桦木科(Betulaceae)鹅耳枥属落叶乔木,花单性,雌雄同株,柔荑花序,雄花序着生于去年生枝上,每苞片具3~13雄蕊,雌花序着生枝顶,每苞片具2雌花,有花被。果序长4~8 cm,具果苞4~8对;小坚果卵圆形,有6~9条中肋及柔毛和腺体。仅分布于浙江省舟山市普陀山岛,1930年由植物学家钟观光教授发现,1932年由林学家郑万钧教授定名。20世纪50年代初,普陀山曾数处发现有该树种分布,但在50年代末,由于大规模的毁林开垦,导致该树种种群规模骤减,仅在佛顶山慧济寺西侧幸存1株母树(卢小根等,1990)。现该母树树高为12.8 m,树干为二分杈,主干胸径63.7 cm,侧干胸径35.3 cm,冠幅12.9 m×11.5 m,树龄约200年。世界自然保护联盟(IUCN)已将该树种列为“严重濒危灭绝(CR)”等级(俞慈英,2007)。1999年经国务院批准,该树种被列为国家Ⅰ级重点保护野生植物(于永福,1999)。为尽快缓解普陀鹅耳枥的濒危现状,保护遗传资源,浙江省舟山市林业科学研究所等单位对该树种的生物学特性与繁殖、栽培、迁地保存技术等进行了一系列的研究攻关,掌握了一套成熟的种质保存技术,自2000年以来,已累计繁育普陀鹅耳枥子代苗木11 896株,有效缓解了其濒危状态。本文对其生物学特性方面的相关繁殖机制的研究结果进行报道。
1 分布区自然概况舟山群岛位于长江以南、杭州湾外缘的东海海域(29°32′—31°04′ N,121°31′—123°25′E)。气候属北亚热带南缘海洋性季风气候区,年平均气温15.6~16.6 ℃,极端最高气温40.2 ℃,极端最低气温-7.9 ℃;年平均降水量936.3~1 330.2 mm,年平均蒸发量1 208.7~1 446.2 mm;年平均日照时数为1 941~2 257 h,年平均风速为3.3~7.2 m·s-1,无霜期251~303天。由于受季风不稳定性影响,夏季易受热带风暴(台风)侵袭,冬季多大风,7—8月间常遇干旱(俞慈英等,2008)。海岛植被属中亚热带常绿阔叶林北部地带的浙闽甜槠(Castanopsis eyrei)-木荷(Schima superba)林区(Ⅳ Aiia-2)(吴征镒,1980)。普陀鹅耳枥母树分布区土壤为黄泥土(石质重壤),母岩为花岗岩,pH5,有机质含量40.4 g·kg-1,全N 2.39 g·kg-1,速效P 2 mg·kg-1,速效N 150 mg·kg-1,土层深厚(卢小根等,1990)。
2 材料与方法以普陀山原生母树和舟山海岛引种驯化树木园(以下简称树木园)内栽培的21年生F1代普陀鹅耳枥植株为对象,开展花期物候观察和花序分布特征、花粉活力、种子品质等生物学特性与繁殖机制研究。
2.1 物候期从2008年2月初开始,每2~3天至树木园内观察普陀鹅耳枥叶、雌花、雄花的物候,记载花芽膨大期、开花始期、开花盛期、开花末期、雄花散粉期、叶芽膨大期、展叶初期、展叶盛期、叶展全期等物候期。
2.2 花序分布特征于2008年4月1日在树木园内随机选定普陀鹅耳枥植株3株,分树冠上、中、下3个部位,按东、南、西、北4个方向各剪取花枝1枝,按上、中、下3个部位分别统计雌、雄花序的数量,分析雌、雄花在树冠上的着生特点和雌雄花序比例。
2.3 花粉活力以树木园内栽培的21年生F1代普陀鹅耳枥植株为对象,于2008年4月1日随机剪取刚开放的雄花花枝若干,带回实验室中水养观察花粉散粉情况,在开花盛期用抖落的方式采集花粉,去除杂质,混匀。将所得花粉分成3份: 1份立即用来培养观察开花盛期的花粉活力;1份在室温22 ℃条件下放置于阴凉干燥处,用来逐日培养观察室温条件下花粉的存活时间;另1份先检测其初始花粉活力, 然后在4 ℃冰箱内干燥贮藏5天后再逐日培养观察花粉活力,用以研究花粉在低温条件下的保存时间。
采用离体萌发法检测花粉活力(黄家兴等,2008;黄修梅等,2008),培养基成分为:0.001%硼酸、10%蔗糖、1%琼脂。具体操作方法:把培养液均匀滴加于载玻片上,在表面形成2~4 mm的薄层。用毛笔蘸取少量花粉,轻轻撒在培养基表面,然后将载玻片放置在垫有湿润滤纸的培养皿中培养,并用另一个培养皿盖住保湿,重复7次。新鲜花粉在培养2 h后用显微镜(OLYMPUS BX50)进行第1次观察,随机镜检花粉萌发情况,4 h后再进行第2次镜检,以后每天镜检2次(10:00,14:00),直至培养的花粉全部死亡为止。室温及4 ℃条件下保存的花粉在培养的次日进行镜检,10:00,14:00各1次,至观察到的花粉全部死亡后停止培养镜检。镜检时,以观察的视野内花粉呈分散状为宜,每次处理计数3~5个视野中的花粉总数和萌发花粉数,统计萌发率。花粉萌发的标准为:花粉管的长度超过花粉半径的1倍视为萌发。判断花粉死亡的标准为:萌发停止, 花粉管萎缩, 花粉孢子解体。
2.4 柱头寿命及可授性以树木园内栽培的21年生F1代普陀鹅耳枥植株为对象,开花后即用放大镜每天观察雌花柱头在色泽、新鲜度等方面的变化情况,以柱头枯萎为衰老特征(齐莉等,2007;叶要妹等,2007),同时检测柱头的可授性。柱头可授性的检测方法为:采集同一时间开放的新鲜雌花用水培养,采取其中的一些柱头,浸在加有联苯胺-过氧化氢反应液(1%联苯胺:3%过氧化氢:水的体积比为4:11:22)凹面载玻片的凹面处,30 min后用放大镜观察柱头的染色情况,如柱头有2/3部位呈现蓝色并有大量气泡产生可认为柱头具有可授性(黄修梅等,2008;齐莉等,2007;叶要妹等,2007;Dafni et al., 1998;彭伟秀等,2008)。
2.5 种子品质2006年12月上旬,对原生母树及F1代所有结果母树进行逐株采种,所采种子进行分株包装、处理与贮藏,测定每株单株采种母树种子的出籽率、千粒质量;分别用解剖法和水选法(下沉种子视为饱满种子,上浮种子视为非饱满种子)测定每株采种母树种子的饱满度。
种子发芽率试验:1)实验室发芽率2007年3月15日,随机取普1号母树及树木园林冠下地面捡拾的混合种子各100粒,消毒后各分成4组置于培养皿内,放在温度为25 ℃、湿度为75%的恒温箱内培养,逐日观察统计发芽种子粒数,计算发芽率。2)场圃发芽率场圃发芽试验包含2个内容:一是在2004年12月,将从树木园林冠下地面捡拾的混合种子,分别用河砂、膨化珍珠岩作贮藏基质,按室内与室外2种摆放环境进行贮藏,2005年3月进行露地播种,2005年6月统计不同方法贮藏的种子场圃发芽率;二是在2006年12月,将各单株母树种子用膨化珍珠岩作基质进行室内贮藏,2007年3月分别进行露地播种,2007年6月统计各单株采种母树种子的场圃发芽率。
2.6 叶芽与枝条萌发力2008年,对容器内栽培的2年生F2代普陀鹅耳枥苗木,春季发芽前抹去枝干中部部分叶芽,观察其隐芽萌发情况。通过控水方法,促使苗木叶片枯萎脱落,顶芽或嫩梢枯死,然后重新供给水分,观察叶芽隐芽与侧芽萌发情况。
2005—2006年,以树木园内栽培的5株18~19年生F1代普陀鹅耳枥植株为研究对象,在植株迁移至浙江林学院植物园时截去树冠上部枝梢,定植后观察被截枝梢的抽发新枝能力。
2.7 幼苗生长特性以普1号母树种子育成的1年生播种苗为研究对象,2007年6月11日至11月15日逐旬定株测定苗高生长量,分析幼苗苗高生长节律,总结幼苗生长特点和栽培管理技术。
3 结果与分析 3.1 开花特性 3.1.1 花期物候观察结果表明,普陀鹅耳枥的雄花先于叶芽和雌花花芽膨大,2月下旬至3月上旬为雄花花芽膨大期,3月中旬进入始花期,3月下旬为开花盛期,4月上旬为开花末期。3月底至4月上旬雄花花药陆续成熟,进入自然散粉期,散粉期持续时间约15天,而单朵花散粉期持续时间约为6天。雌花花芽在3月中旬至3月下旬膨大,3月底至4月初为雌花始期,4月上旬进入开花盛期,4月中旬为开花末期。雌、雄花相遇期约16天,比卢小根等(1990)报道的相遇时间9天有明显延长。叶芽膨大期与雌花花芽膨大期一致,3月底为展叶初期,4月上旬为展叶盛期,4月中旬前后为叶全展期。
3.1.2 花序分布特征统计结果(表 1)表明,普陀鹅耳枥的雄花序在树冠上部着生的占34.93%,树冠中部占36.90%,下部占28.17%,三者基本呈均匀分布。这个统计结果与卢小根等(1990)报道结果(下部占50%,中部占33.5%,顶部仅占16.7%)有较大出入。而雌花序的垂直分异明显,以树冠上部占绝对优势,达52.34%,中部占36.72%,上、中部之和达到了89.06%,而树冠下部仅占10.94%。这个统计结果与卢小根等(1990)报道结果相吻合。
|
|
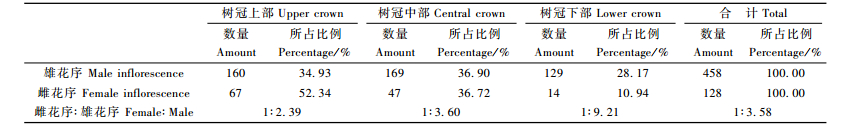
从雌、雄花序数量比例来看,总体上比例为1:3.58,与卢小根等(1990)报道结果14.5%:85.5%(1:5.90)有差异。从树冠上、中、下3个部位来看,上部雌、雄花序的比例为1:2.39,中部为1:3.60,下部为1:9.21,下部雌花序所占比例显著低于中、上部。
3.2 花粉活力 3.2.1 开花盛期的花粉活力试验结果(图 1)表明,在开花盛期采集的花粉,萌发率最高峰出现在培养的第2天与第3天,平均最高萌发率达到了52.41%,比卢小根等(1990)研究结果(最高萌发率为29%)要高,但总体萌发率仍较低。第4天开始萌发率下降,至第6天观察时,仅有极少数孢子还有活力,至第7天观察时,孢子均开始裂解死亡。

|
图 1 普陀鹅耳枥花粉萌发率检测结果 Figure 1 Pollen germination percentages of Carpinus putoensis 萌发率均为7组重复数据的平均数Germination percentages are the mean of the 7 repeats. |
试验结果(图 1)表明,普陀鹅耳枥花粉在室温22 ℃条件下干燥贮藏的存活寿命在6天左右,略低于卢小根等(1990)报道的“在常温下能保存7~12天”的研究结果。花粉在培养的第3天萌发率最高,平均萌发率达到51.01%,以后随着贮藏时间的增加,其萌发率开始下降,在贮藏6天后花粉孢子死亡。
3.2.3 4 ℃条件下贮存的花粉活力试验结果(图 1)表明,放置于4 ℃冰箱内干燥保存的花粉起初萌发率最高在48.27%左右,低温保存5天后,花粉萌发率开始下降。从开始观察起到孢子死亡共为15天,即低温贮藏的花粉可以延迟花粉活力10天左右且萌发率与室温条件下保存的萌发率结果基本相同,这说明低温干燥条件下贮藏可以较好地延长普陀鹅耳枥花粉的活力,从而可以在一定程度上缓解雌雄花花期相遇时间短所带来的授粉困难问题,为人工辅助授粉提供了依据。
3.3 柱头寿命及可授性检测观察结果表明,雌花开放初期,柱头呈新鲜的粉红色,到第3天时,部分柱头开始成浅褐色,新鲜度显著降低,略成枯萎状;到开放的第5天,柱头则全部变得干枯萎蔫。柱头可授性检测结果(表 2)表明,雌花从开放开始柱头即具有可授性,在开放后24 h至3天,柱头的可授性最强,至开放的第4天开始,柱头的可授性迅速下降,至第5天便完全丧失了可授性,前后持续时间仅4天。柱头可授性变化与其自身的颜色和新鲜度的变化存在一定的相关性,故可以通过放大镜观察柱头的色泽来辅助判断柱头的可授性。
|
|
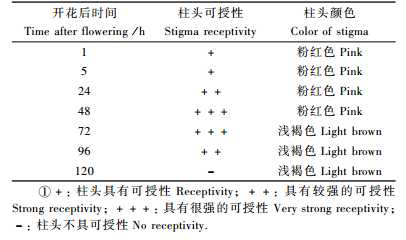
柱头可授性结合花期物候观察结果表明,4月上旬至中旬为柱头可授期,持续时间约14天,单个柱头可授期持续时间约为4天,但雌花柱头可授期与雄花花粉散粉期重叠时间仅为6天,说明普陀鹅耳枥的有效授粉时间很短。
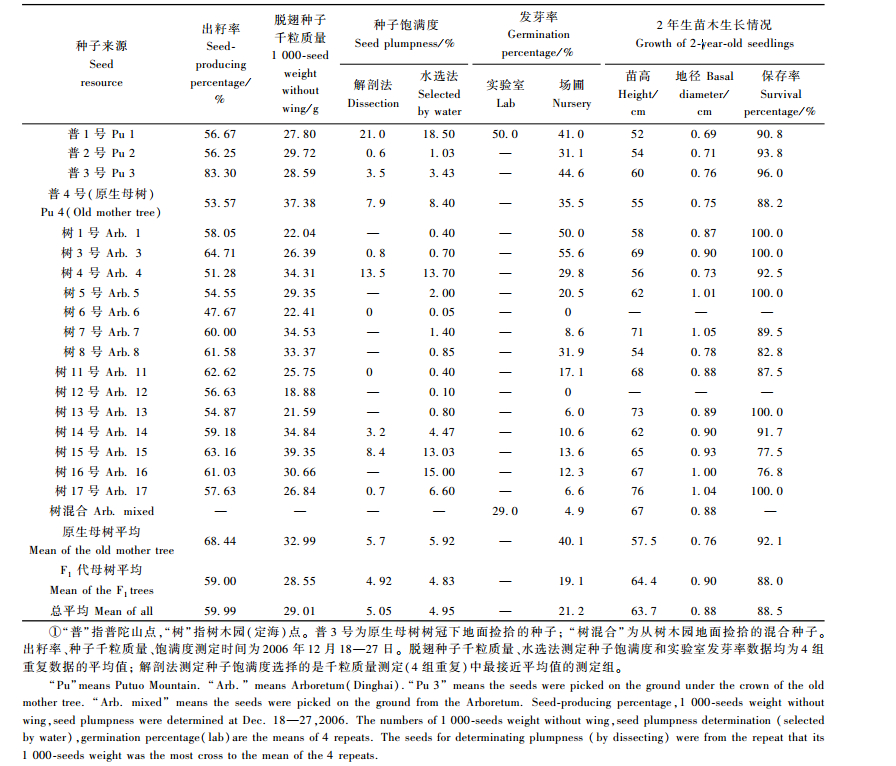
3.4 种子特性 3.4.1 种子品质检验结果(表 3)表明,不同母树上采集的种子,其出籽率、千粒质量、饱满度等品质指标均有较大差异,平均出籽率为59.99%,平均脱翅千粒质量为29.01 g。解剖法测定的种子平均饱满度为5.05%,水选法测定的种子平均饱满度为4.95%,从总体上看,2种方法测定的饱满度比较接近,所以在生产应用中,可以用水选法代替解剖法来测定普陀鹅耳枥种子的饱满度,避免解剖造成种子浪费。
|
|
实验室发芽率试验结果表明,各小组均在培养2天后(3月17日)开始发芽,普1号各组种子在8~12天基本停止发芽,有2组分别在第24天和第28天还有个别种子继续发芽,平均发芽率为50.0%;树木园混合种子在培养4~11天后即陆续停止发芽,平均发芽率为29.0%。
场圃发芽率试验结果(表 3)表明,不同母树上采集的种子,其场圃发芽率有明显的差异,平均发芽率为21.2%,其中,原生母树种子的平均场圃发芽率为40.1%,F1代母树种子的平均发芽率为19.1%,原生母树种子的发芽率要显著高于F1代母树种子。
另外,从总体平均来看,原生母树种子的各项品质指标均要高于F1代母树所产种子,说明原生母树的种子品质要优于F1代所产种子,这可能与F1代母树年龄较小,尚未进入结实盛年期有关。在F1代母树中,普1号所产种子的品质要显著优于其他F1代母树所产种子,这可能与其处于片林中间、授粉几率高,与周边树木间距较大、光合作用充分等原因有关。
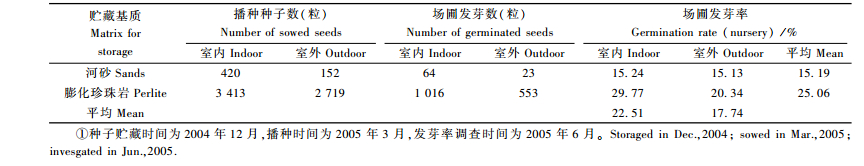
种子贮藏试验结果(表 4)表明,用膨化珍珠岩贮藏的种子,其场圃发芽率要明显高于用河砂贮藏的种子,这可能与膨化珍珠岩透气性好、无菌的特点有关。室内贮藏的种子场圃发芽率要高于室外贮藏的种子,这主要与室内贮藏管理精细有关。
|
|
试验结果表明,普陀鹅耳枥2年生苗枝干中部的叶芽被抹除后,其隐芽将不再萌发。在控水条件下,叶片枯萎脱落后再重新给水,也未见隐芽萌发,但顶芽(包括侧枝的顶芽)或嫩梢枯死后,在重新给水养护情况下,枯死顶芽(或嫩梢)以下的1至多个活侧芽能重新萌发抽梢。迁移至浙江林学院植物园的5株18~19年生植株,其被截枝干均重新抽发了新梢,这说明普陀鹅耳枥具备一定的萌芽更新能力,与前人研究结论(萌芽性很差)(卢小根等,1990)有差异。
3.6 幼苗生长特性普陀鹅耳枥种子播种后,一般在4月下旬出苗,7月上旬开始幼苗高生长进入速生期,7月中旬至8月下旬为幼苗高速生期,其中8月中旬为高生长高峰期,9月上旬开始幼苗生长减缓,9月下旬开始停止生长,全年平均苗高生长量为37.7 cm(图 2)。

|
图 2 普陀鹅耳枥1年生幼苗高生长节律 Figure 2 Height growth of one-year-old seedlings of C. putoensis |
研究表明,普陀鹅耳枥苗期应进行多次移栽,以促进其根系及地径生长。同时应及时做好拔草、施肥、松土等工作。幼苗易遭地下害虫为害,可及时用1:2 500倍锌硫磷溶液防治。幼苗生长喜土壤深厚、排水良好、具侧方庇荫的条件。不同母树育成的2年生播种苗平均高生长量为52~76 cm,平均地径生长量为0.69~1.05 cm,苗木保存率为76.8%~100.0%(表 3)。
4 结论与讨论 4.1 普陀鹅耳枥的繁殖机制是导致其濒危的生物学原因植物的繁殖包括有性繁殖和无性繁殖。种子植物在地球上繁盛并占尽优势的一个重要原因是其花器官及种子的繁殖。由合子发育的幼胚与包藏幼胚的种皮构成的种子,既是植物种群杂交系统给自然选择提供的遗传变异组合极丰富的素材,又是植物生活史中对环境压力具有高度耐受性和应变力的一个阶段(孙凡等,1997)。植物生活史中以种子形态出现的阶段被称为植物的潜在种群(谢宗强等,2000;Harper,1977;Silvertown,1982),以有性繁殖为主的植物,其种子产量、天然条件下发芽率、成苗定居率是生活史中最关键的环节,关系到种群能否实现更新与维持(张文辉等,2006),因此,种子对于植物种群的分布格局、种群动态及种群调控具有重要作用,能否有足够的种子种群经散布后形成土壤种子库,避开不良环境,在适宜时机达到幼苗的建成,为种群补充后代,对种群的发展和壮大至关重要(Harper,1977;郑宏春等,2004;谢宗强等,2000;陈远征等,2006)。就普陀鹅耳枥而言,虽然在扦插、嫁接、组培等无性繁殖技术上已取得了重要突破,为尽快扩大种群规模提供了支撑,但从有效保存其遗传基因的多样性和持续进化潜力,促进野外种群的自我重建角度考虑,研究其有性繁殖机制,提高有效结实量和种子生活力显得尤为迫切。
本研究表明,普陀鹅耳枥的繁殖机制是导致濒危的生物学原因,其有性繁殖的主要障碍在于其有效结实量少、种子生活力差、品质低劣,而且结实的大小年现象明显,而影响其有性繁殖有效性的原因主要有:
1) 授粉几率低 一方面,植物授粉与雌雄花的花期重叠密切相关。普陀鹅耳枥的雄花先于雌花开放,雌雄花花期相遇期约为16天,但雄花散粉期与雌花柱头可授期重叠时间仅为6天左右,这种错开式的花期使得雌花的授粉几率大大降低,严重影响了种子的饱满度和萌发率,也影响了该物种的天然更新。另一方面,在植物繁殖过程中,雌雄花的着生部位和数量比例与传粉有着十分密切的关系。观察结果表明,普陀鹅耳枥雄花数量是雌花的3.58倍,这保证了雄花在传粉时能散发大量的花粉来确保授粉的完成,但从花的着生部位来看,普陀鹅耳枥的雄花在树冠上、中、下部基本呈均匀分布,但雌花则集中分布于树冠的上部和中部,这样的着生方式非常不利于授粉,制约了传粉的有效性,从而影响了种子的产量和质量。
2) 花粉活力低 花粉作为植物的雄配子体,在有性生殖中发挥着重要的作用,花粉发育及花粉管的萌发和生长是植物有性生殖过程中的重要事件(孙春丽等,2008),花粉活力和萌芽率是影响受精能否成功的最关键因素。本试验结果表明,普陀鹅耳枥花粉的总体萌发率不高,加上存活时间很短,进一步导致了授粉的成功率降低。
3) 传粉与授粉受花期恶劣气象条件影响较大 传粉是种子植物种群生活史中一个重要的生命活动现象,是种子植物受精的必经阶段;花粉的运动在很大程度上决定了植物个体间的基因流和群体的交配方式,对种群的遗传与变异起着重要作用(鲁先文等,2008)。普陀鹅耳枥的花序为柔荑花序,为风媒植物特有花序,花粉依靠风力传播,花粉的释放、散布对风、温度、降雨等气象条件的要求较高(马缘生,1989)。风媒植物的传粉有效性通常较虫媒植物低(林树燕等,2008)。普陀鹅耳枥花期在每年的3月中旬开始持续到4月中旬,这个时期也正是舟山一带阴雨天气较多的时候,降雨和大风使得雄花散粉和花粉传播受到巨大的阻碍,并造成大量落花,给雌花受孕带来了较大的困难,从而严重影响了种子的产量和品质,给有性繁殖后代带来了困难。
4.2 基因资源保存措施从普陀鹅耳枥繁殖机制及致濒原因研究结果可知,如何提高普陀鹅耳枥种子的产量和品质,尽快扩大有性后代的种群数量,并逐步恢复其天然更新能力,将是解除其濒危状态,有效保护物种基因资源的根本途径。可以通过一些人工辅助手段来提高其有性繁殖能力:
1) 花粉低温储藏 花粉的长期贮藏对种质保存和杂交育种十分重要(王彩虹等,1996),为解决亲本花期不遇和远距离杂交等问题,更需要保存有生活力的花粉(王红等,2006)。通过低温冷藏的花粉可以引起花药内小孢子的营养饥饿,启动营养细胞DNA的复制、分裂和进一步发育,从而提高小孢子的生活力,达到延长花粉寿命的目的(李文泽等,1995)。本研究也表明,采集普陀鹅耳枥盛花期的花粉,在4 ℃低温条件下进行干燥冷藏能有效延长花粉的活力,可在一定程度上缓解雌雄花花期相遇时间短而造成的授粉困难问题。另据报道,采用超低温技术保藏的花粉,在保存1年时间后解冻复苏仍可较好地恢复生活力(马缘生,1989;王彩虹等,1996;王红等,2006),此种方法能否用于普陀鹅耳枥的花粉保存值得研究。
2) 人工辅助授粉 人工辅助授粉授粉可以克服风媒传粉受花期天气限制大(吴小明等,2008)、雌雄花序着生部位不合理等影响传粉、授粉的重大关键问题,因此能很好地解决普陀鹅耳枥自然授粉困难这一问题,有望成为提高普陀鹅耳枥种子品质,从而提高有性繁殖系数的有效手段之一。
3) 加强营林措施 分析本研究与卢小根等(1990)在雌雄花相遇期、花粉活力等方面研究结果存在的差异,可能与两者的研究对象所处环境与群体大小等原因有关。卢小根等(1990)以单株的原生母树为研究对象,该树生长于普陀山佛顶山西侧,花期受大风等恶劣天气影响较大,易造成雄花早落,同时因花粉源少且传粉受大风等天气影响大而降低了传粉的有效性,从而导致了雌雄花相遇期缩短、花粉活力不高的结果。而本研究以位于定海树木园内的F1代子代林为研究对象,该子代林所处位置为沟谷区域,受大风等影响较小,个体数量较多(共17株),利于雄花发育、散粉和传粉,所以雌雄花相遇期较原生母树显著延长,雌花授粉较充分,花粉活力明显提高。因此,营林措施在普陀鹅耳枥基因资源保存中具有重要的作用。首先,要根据普陀鹅耳枥的生物学、生态学特性,选择土壤肥沃、排水良好的沟谷区域营建子代林,切勿在迎风的山岗等区域造林,以促进植株健康生长,并可避免花、果期遭遇大风而引起落花落果。其次,普陀鹅耳枥为阳性树种,在造林时必须合理控制林分密度,保证结实母树能得到充足的光照,有效提高光合作用效能,同时做好水、肥、病虫害等其他抚育管理措施,以提高母树的结实能力,促进结实。第三,营造的林分应达到一定规模的群体数量,以增加相互之间的授粉几率,提高种子的产量和品质。
陈远征, 马祥庆, 冯丽贞, 等. 2006. 濒危植物沉水樟的濒危机制研究[J]. 西北植物学报, 26(7): 1401-1406. |
黄家兴, 吴杰, 安建东, 等. 2008. 凯特杏花粉的离体培养及影响因子分析[J]. 西北植物学报, 28(1): 52-58. |
黄修梅, 郝丽珍, 胡宁宝, 等. 2008. 沙芥花粉萌发特性和柱头可授性的研究[J]. 园艺学报, 35(10): 1473-1478. DOI:10.3321/j.issn:0513-353X.2008.10.011 |
李文泽, 胡含. 1995. 在花药花粉培养中预处理的作用机理[J]. 遗传, 17(增刊): 13-18. |
李忠超, 王武源. 2006. 濒危药用植物八角莲生态生物学特性[J]. 热带亚热带植物学报, 14(3): 190-195. |
林树燕, 丁雨龙, 张昊. 2008. 5种竹子花粉萌发率及开花特性[J]. 林业科学, 44(10): 159-163. DOI:10.3321/j.issn:1001-7488.2008.10.027 |
鲁先文, 马瑞君, 孙坤. 2008. 中国沙棘(Hippophae rhamnoides L. ssp. sinensis Rousi)的开花特性及风媒传粉距离的检测[J]. 生态学报, 28(6): 2518-2525. |
卢小根, 邹达明. 1990. 普陀鹅耳枥濒危原因的调查研究[J]. 浙江林业科技, 10(5): 61-64. |
马缘生. 1989. 作物种质资源保存技术[M]. 北京: 学术书刊出版社: 200-217.
|
彭伟秀, 冯大领, 郑辉, 等. 2008. 晚熟李花粉生活力及柱头可授性研究[J]. 河北农业大学学报, 31(5): 57-60. |
齐莉, 巨艳秋, 李微, 等. 2007. 梨花柱头可授性和花粉活力的研究[J]. 牡丹江师范学院学报:自然科学版, 60(4): 29-30. |
孙春丽, 潘延云. 2008. 拟南芥花粉活力的测定及其在花粉发育研究中的应用[J]. 植物学通报, 25(3): 268-275. |
孙凡, 钟章成. 1997. 植物繁殖生态学:进化生态学与行为生态学的结合点[J]. 科学(中文版), (8): 59-61. |
王彩虹, 李嘉瑞. 1996. 杏花粉的低温与超低温贮藏研究[J]. 莱阳农学院学报, 13(2): 169-173. |
王红, 王春梅, 黄鲲. 2006. 柑桔花粉超低温保存初探[J]. 中国南方果树, 35(4): 8-10. |
吴小明, 葛世魁, 方益柱, 等. 2008. 南方红豆杉幼树人工授粉技术研究[J]. 江西农业学报, 20(12): 50-51. DOI:10.3969/j.issn.1001-8581.2008.12.017 |
吴征镒. 1980. 中国植被[M]. 北京: 科学出版社: 851-853.
|
谢宗强, 李庆梅. 2000. 濒危植物银杉种子特征的研究[J]. 植物生态学报, 24(1): 82-86. |
叶要妹, 张俊卫, 齐迎春, 等. 2007. 百日草柱头可授性和花粉生活力的研究[J]. 中国农业科学, 40(10): 2376-2381. DOI:10.3321/j.issn:0578-1752.2007.10.036 |
俞慈英. 2007. 普陀鹅耳枥[J]. 科学世界, (6): 56. |
俞慈英, 李修鹏, 赵慈良, 等. 2008. 普陀樟生物学特性与栽培技术[J]. 林业科学, 44(9): 65-71. DOI:10.11707/j.1001-7488.20080912 |
于永福. 1999. 中国野生植物保护里程碑——国家重点野生植物名录(第一批)[J]. 植物杂志, (5): 3-11. |
张文辉, 许晓波, 周建云. 2006. 濒危植物秦岭冷杉生殖生态学特性[J]. 生态学报, 26(8): 2417-2424. |
郑宏春, 路安民, 胡正海. 2004. 数珠珊瑚(桑陆科)的花器官发生[J]. 西北植物学报, 24(3): 476-483. |
Dafni A, Maués M M. 1998. A rapid and simple procedure to determine stigma receptivity[J]. Sexual Plant Reproduction, 11: 177-180. DOI:10.1007/s004970050138 |
Harper J L. 1977. Population biology of plants[M]. London: Academic Press.
|
Lande R. 1988. Genetics and demography in biological conservation[J]. Science, 241: 1455-1460. DOI:10.1126/science.3420403 |
Silvertown J W. 1982. Introduction to plant population ecology[M]. London: Longman: 92.
|