 2010, Vol. 46
2010, Vol. 46文章信息
- 罗大庆, 王军辉, 任毅华, 朱登强
- Luo Daqing, Wang Junhui, Ren Yihua, Zhu Dengqiang
- 西藏色季拉山东坡急尖长苞冷杉林的结实特性
- Fruiting Characteristics of Abies georgei var. smithii Forest on the Eastern Slope of the Sejila Mountain in Tibet
- 林业科学, 2010, 46(7): 30-35.
- Scientia Silvae Sinicae, 2010, 46(7): 30-35.
-
文章历史
- 收稿日期:2009-02-17
- 修回日期:2009-10-12
-
作者相关文章


2. 中国林业科学研究院林业研究所 北京 100091
2. Research Institute of Forestry, Chinese Academy of Forestry Beijing 100091
植物的球果和种子是物种繁殖系统的重要特征,是植物种群生殖生态学研究的基础,由于处于强大的选择压力之下,因而表现出对环境的较大适应性(Wheeler et al., 1982)。同时,球果和种子也是受遗传控制较强的特征,最具有区分和比较的意义(Ruby,1967; 孙玉玲等,2005)。在植物的生活史中种子是一个重要的阶段,它关系到种群的未来命运,种子阶段是有性繁殖的植物个体一生中唯一有移动力的阶段,对于植物种群的分布、动态及调控等方面均有重要意义(Harper,1977; Steven,1991; 谢宗强等,1998)。开展种实研究对于物种的群落演替和种群更新,及其生态适应过程和系统发育过程都具有重要意义(徐亮等,2004)。
急尖长苞冷杉(Abies georgei var.smithii)是松科冷杉属常绿乔木,主要分布于滇西北、川西南和西藏东南部地区。它是冷杉属在西藏分布面积最广的一个树种,通常在西藏东南部受季风影响的高山峡谷区上部形成暗针叶林,成为湿润山地亚高山地带的显域植被和顶级群落; 暗针叶林在西藏的分布和盛衰,与青藏高原的隆起和地质时期的冷暖变化有着密切的关系,它是长期的历史过程中,生物与气候相互作用的产物(李文华,1985),在维持高海拔地区的生物多样性与生态平衡方面具有不可替代的作用。有关急尖长苞冷杉种群、群落以及生态系统层次的生态学研究在西藏高山森林生态系统定位研究中已研究报道(任青山,2002; 辛学兵等,2003; 罗大庆等,2003),然而关于球果以及种子形态特征及其生境的研究,目前还未见报道。研究急尖长苞冷杉的结实特性,有助于阐明该树种的繁殖适应性,对分析冷杉种群生存力和种群对策至关重要。因此,本研究拟通过对急尖长苞冷杉林结实特性的调查,分析探讨其基本特征与海拔生境的关系,为这一树种的合理利用和人工培育提供理论依据,同时也是西藏高山森林生态系统长期观测研究必须具备的基础数据。
1 研究区自然概况与研究方法 1.1 自然概况研究区域位于西藏东南部的林芝地区林芝县境内色季拉山东坡。色季拉山是念青唐古拉山向南延伸的余脉,它与喜马拉雅山脉东端向北发展的山系相邻,与南迦巴瓦峰相望。色季拉山体海拔在2 100~5 300 m之间,水平走向大致呈西北向东南,由此形成了较大范围的东、西坡面。东坡承雅鲁藏布江水汽通道作用,南来的印度洋暖湿气流沿帕龙藏布河谷进入本区,带来丰沛的降水; 该区气候湿润,属亚高山寒温带湿润气候区。根据东坡海拔3 850 m处定位站2001—2007年的气象观测数据显示,该区年平均气温2.0~4.5 ℃,最暖月(7月)平均气温11.1 ℃,最冷月(1月)平均气温-4.1 ℃; 年降水量875.5~1 350.9 mm,平均1 095.4 mm,主要集中在4—9月,占全年降水的88%;年蒸发量601.7 mm,年均相对湿度83%。
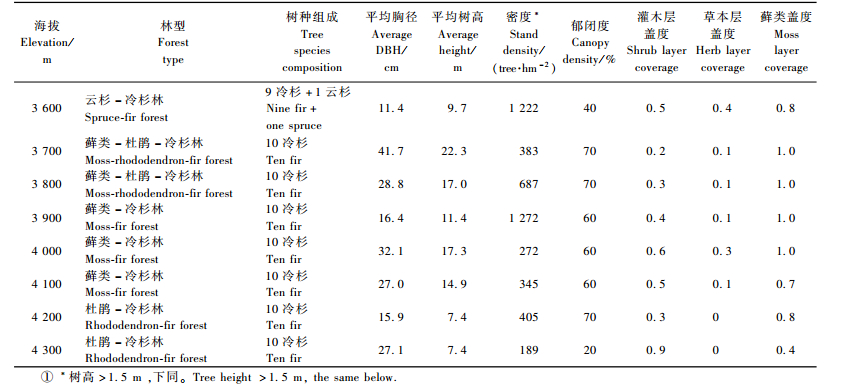
急尖长苞冷杉林在色季拉山东坡分布的基海拔大致在3 550 m左右,最高至林线4 300 m(树木线4 350 m),属保存完整的原始林,其中,海拔3 600~4 000 m为最适分布地带,自然长势最好,本次调查的林分主要特征如表 1所示,平均林龄大约在200年左右,属成过熟林,优势木胸径118.0 cm, 树高可达50.7 m; 乔木层组成树种单一,林地藓类极度发育,盖度在0.4~1.0之间。其他树种有林芝云杉(Picea likiangensis var.linzhiensis)、西南花楸(Sorbus rehderiana)、方枝柏(Sabina saltuaria)、高山柏(Sabina squamata); 群落灌木层植物由多种杜鹃(Rhododendron spp.)、峨嵋蔷薇(Rosa omeiensis)、陇塞忍冬(Lonicera tangutica)、青藏垫柳(Salix lindleyana)、刺茶藨子(Ribes alpestre)、冰川茶藨子(Ribes glaciale)组成; 草本层主要以西南草莓(Fragaria moupinensis)、凉山悬钩子(Rubus fockeanus)、糙野青茅(Deyeuxia scabrescens)、紫鳞苔草(Carex souliei)、蓼(Polygonu spp.)、五裂蟹甲草(Cacalia pentaloba)、锡金柳叶菜(Epilobium sikkimense)、白花酢浆草(Oxalia acetosella)、大叶碎米荠(Cardamine macriphylla)、草莓凤仙花(Impatiens fragicolor)、光茎四川堇菜(Viola szetchwanensis var.nudicaulis)、小舌紫菀(Aster albescens)、鞭打绣球(Hemiphragma heterophyllum)、堇花唐松草(Thalictrum diffusiflorum)等植物为主; 藤本植物有防已叶菝葜(Smilax menispermoides); 群落中蕨类植物有尖齿鳞毛蕨(Dryopteris acutodentata)、西藏铁线蕨(Adiantum tibeticum)、巢蕨(Neottopteris spp.)等。
|
|
从2000—2008年连续观测急尖长苞冷杉的结实大小年变化。在2006年(种子丰年)10月中旬,急尖长苞冷杉球果基本成熟,种鳞未张开,种子尚未散落前开始采集球果。采集样地选在西藏高原生态研究所色季拉山森林生态定位研究站的急尖长苞冷杉林垂直样带固定样地中,样带从海拔3 600 m至林线4 320 m,沿垂直等高线方向每间隔100 m设置1个30 m×30 m的固定样地,同一坡向上共设8个固定样地。在每个固定样地中进行每木调查,并统计结实母树的数量和比例,算出结实母树的平均胸径和树高,选取3株接近于平均值的结实母树作为球果采集的标准株。将标准株树冠等分成上、中、下3层,每层按东、西、南、北4个方位计数球果数量并采集球果样本,球果样本按大、中、小3个等级各采集3~5个,分别编号装入布袋带回实验室。球果形态特征的测量是在球果开裂之前,用游标卡尺实测每个球果样本的长度、宽度; 待球果干燥开裂,种子和种鳞自然脱落后计数每球果样本的种子数量,从每个方位的样本中随机选取30粒种子用游标卡尺量测种子长度、宽度、厚度; 用百粒法测定千粒质量,将每个标准株的球果样本种子净种后,用四分法随机从每个样本中分别取出100粒为1组,用1/1 000电子天平称取质量并推算千粒质量,通过解剖种子,人为目视鉴别胚的完好程度,分别计数被测样本种子的饱满度(Li et al., 1997)。数据统计和分析分别用Office Excel和SPSS 13.0软件进行。
2 结果与分析 2.1 结实开始期和结实周期种子的生产是林木生活史中一个重要的阶段,主要反映在林木开始结实的年龄早晚、周期长短和种子大小等方面。对急尖长苞冷杉天然林结实年龄调查发现,其初始平均结实年龄通常在20年左右,该年龄的平均树高9.8 m左右,胸径20.5 cm左右; 进入结实期的成龄树直至老龄阶段均能够持续结实,但不同年龄阶段有所差异,一般幼龄至中龄阶段的结实能力较强。
根据连续观察年份的林分结实情况调查结果发现,急尖长苞冷杉天然林通常2~3年结实1次。所观察的年份中,2003年和2006年为种子大年,林木普遍结实,且结实量大,其间隔的2年中,第1年不结实,第2年有单株少量结实,但结实量较小,即种子大年之后的第2年不结实,随后出现1个结实小年,再进入下一个结实大年。这一现象说明,急尖长苞冷杉的结实具有一定规律,属于有明显间隔周期的格局类型。
2.2 急尖长苞冷杉林结实状况森林群落中,成龄母树的结实比例和数量通常作为衡量林分繁殖能力的主要标准。由于不同海拔梯度上林分的组成、密度和生长状况均有差异,因此会对林分结实产生影响。表 2的统计结果显示,急尖长苞冷杉林的结实母树比例在25%~80%之间,平均为45%,与秦岭冷杉(Abies chensiensis)林55%的结实比例相比(孙玉玲等,2005),急尖长苞冷杉林结实母树比例偏低; 结实母树数量在131~483株·hm-2之间,平均为230株·hm-2; t检验证明,在0.05置信水平上,不同海拔的结实母树比例(Sig.=0.000)和结实母树数量(Sig.=0.001)均有显著差异,说明不同海拔的急尖长冷杉种群繁殖力存在差异; 从结实母树数量的比较结果来看,3 900 m以下各海拔的结实状况明显优于4 000 m以上的其他海拔。
|
|
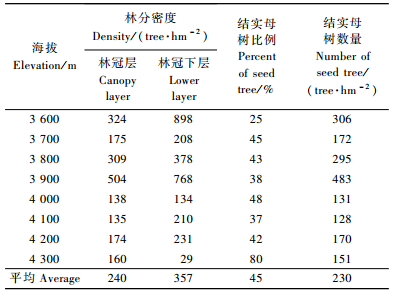
进一步分析结实母树数量特征值与林分密度特征值的关系,结果显示,各海拔的结实母树数量与林分密度以及冠层木数量、下层木数量均有极显著相关,相关系数分别为0.91,0.99,0.84;结实母树比例与林分密度和下层木比例呈显著负相关,与冠层木比例呈极显著正相关,相关系数分别为-0.64,-0.99,0.99。这一分析结果表明,急尖长苞冷杉林的结实母树数量受到林分密度控制,并随着海拔梯度上的林分密度的变化发生改变,其中,冠层木的比例和数量决定了结实母树的比例和数量; 随着海拔升高,林木逐渐稀疏,林分密度下降,结实母树的数量递减,而结实母树比例则有所增加。
2.3 单株结实量与空间分布特征分析不同海拔急尖长苞冷杉林的单株结实情况及其林分生长状况的关系,对各海拔结实母树的平均胸径、树高、冠幅以及球果空间分布数量统计结果如表 3所列。急尖长苞冷杉林单株球果数量24~366个,平均为157个,单株个体以及不同海拔间的结实量差异较大。其中,海拔4 000~4 100 m的平均单株结实量最高,平均295~366个; 冷杉林分布上限海拔4 300 m的单株结实量最低,平均单株结实仅24个; 其余各海拔的单株结实量在60~202个之间; t检验证明,在0.05水平上,不同海拔间的单株结实量差异显著(Sig.=0.008)。随着海拔的升高,结实母树的平均胸径和树高呈递减的趋势,而平均单株结实量的变化无明显规律,相关分析结果表明,急尖长苞冷杉的单株结实量与胸径和树高无显著相关。
|
|
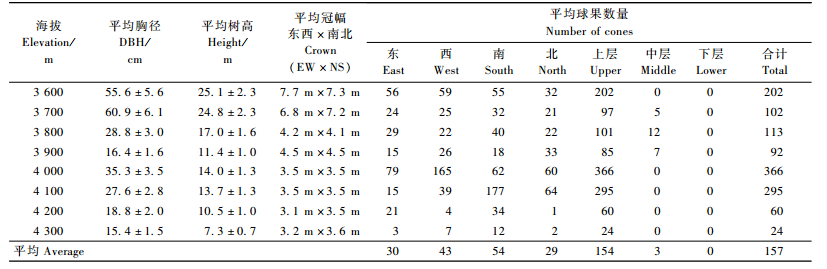
从急尖长苞冷杉结实的空间分布来看,垂直空间上,98%的球果集中分布于树冠上层,其余2%分布于中层,下层无球果分布; 球果的水平分布在不同树冠方位上以南向分布居多,西向次之,东向和北向相对较少但相差不大。急尖长苞冷杉的球果空间分布特征与秦岭冷杉和元宝山冷杉(Abies yuanbaoshanensis)的基本一致(唐润琴等, 2001; 孙玉玲等,2005)。
2.4 球果特征由表 4可以看出,不同海拔的急尖长苞冷杉球果各项特征值均存在差异。急尖长苞冷杉球果的长度在2.22~9.82 cm之间,平均6.39 cm±0.2 cm; 球果宽度在2.35~5.42 cm之间,平均3.45 cm±0.1 cm; 各海拔中,以3 800 m和3 900 m的球果最大,4 000, 4 200和4 300 m的球果较小。单个球果中包含的种子数量差异较大,在85~720粒之间,平均441粒±20粒,其中单果包含种子数量较多的海拔是3 600 m和3 800 m,平均超过500粒; 4 100 m以上各海拔的单果种子数量较少,多数少于400粒。各海拔的平均单果种子饱满率在16%~74%之间,差异较大,平均为49.62%±5.5%;其中,冷杉林下限海拔3 600 m的种子饱满率最高,海拔4 200 m的种子饱满率最低。
|
|

比较不同海拔的球果形态,方差分析表明,在0.05置信水平上,球果长度、单果种子数量和饱满率3项指标差异极显著(P<0.01),球果宽度差异不显著(P>0.05)。随着海拔升高,球果的各项特征值的变化较大,总体上呈下降的趋势,表明急尖长苞冷杉林分布的低海拔区的种群生殖状况要优于高海拔。与冷杉属其他树种相比较(孙玉玲等,2005; 赵德明,2001),急尖长苞冷杉的球果长度略短,宽度居中,单果种子数量较高。
2.5 种子特征在植物的诸多性状中,种子大小处于中心地位,是植物生活史中的核心特征(Fenner et al., 2005; Westoby et al., 1992)。对急尖长苞冷杉种子大小和千粒质量4项性状的测定结果(表 5)表明:急尖长苞冷杉的种子长度为0.46~0.86 cm,平均0.67 cm; 宽度为0.13~0.32 cm,平均0.24 cm; 厚度为0.09~0.19 cm,平均0.15 cm; 千粒质量为2.72~17.04 g,平均8.14 g。在冷杉属树种中,急尖长苞冷杉种子较小(孙玉玲等,2005; 赵德明,2001)。用变异系数比较各海拔的种子性状变化,变异系数越大则离散程度越大。比较结果显示,各海拔的种子长度变异系数为5.13%, 宽度变异系数为5.68%, 厚度变异系数为5.19%, 种子千粒质量的变异系数为22.84%;其中,千粒质量的变异最大,种子大小的变异较小,表明急尖长苞冷杉的种子大小是相对稳定性状,而种子质量则变化较大。
|
|
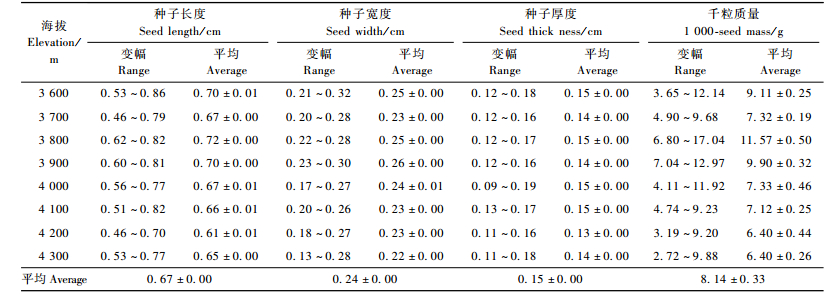
从各海拔的种子性状变化情况来看,图 1显示,种子的长度、宽度、厚度以及千粒质量4项指标随着海拔升高均呈递减的趋势,其中,海拔3 800 m和3 900 m的种子综合性状在各海拔中最优; 总体上,急尖长苞冷杉林分布的低海拔区的种子性状明显优于高海拔,这与前述球果特征的结论是一致的。

|
图 1 不同海拔急尖长苞冷杉种子形态特征比较 Figure 1 Comparison of the seed morphological characteritsics of A. georgei var. smithii at different elevations |
急尖长苞冷杉是藏东南亚高山暗针叶林的主要组成树种之一,在喜马拉雅山脉东段北坡海拔3 600~4 300 m的地带连续分布,并构成该地带显域植被和顶级群落。本文通过其典型分布区色季拉山急尖长苞冷杉林的结实调查与分析,结果表明,急尖长苞冷杉林的结实属于有明显间隔期的格局类型,通常3年出现1次结实丰年,天然林初始结实年龄在20年左右,该时期树高在9.8 m左右,胸径20.5 cm左右。
本研究区的急尖长苞冷杉林垂直分布跨度较大,不同海拔间,由于林分密度的差异,结实母树数量也存在较大差异,结实母树数量在131~483株·hm-2之间; 随着海拔梯度升高,林分逐渐稀疏,结实母树的数量递减,而结实母树比例有所增加。就群体而言,急尖长苞冷杉林的结实表现为林冠上层林木结实,下层和林内不结实; 单株树冠上部结实,下部不结实; 树冠南面结实较多。这一特点表明,急尖长苞冷杉虽属耐荫性树种,但其生殖生长时期仍是一个需光的过程,较好的光照和温度条件对其开花、结实具有积极的意义。
不同海拔急尖长苞冷杉林的单株结实有差异,在垂直分布上无明显规律,单株结实球果从24~366个不等,其中海拔4 000~4 100 m的平均单株结实量最高; 分析结果显示,急尖长苞冷杉的单株结实量与胸径和树高无显著相关。与冷杉属其他树种相比,急尖长苞冷杉的球果长度略短,宽度居中; 种子长度、宽度和厚度均小,单果种子数量较多。不同海拔的比较结果表明,急尖长苞冷杉球果和种子均以低海拔区发育最好,球果和种子较大,千粒质量、出种量以及饱满度均优于高海拔。
已有研究表明,由于冷杉属树种的垂直分布海拔范围较大,而不同海拔间气候、立地条件、群落结构均有所差异,因此球果和种子在形态和遗传上会随之发生变异(孙玉玲等,2005)。本文的分析结果也显示,急尖长苞冷杉的种实也有类似的垂直分布变异,其结果有待深入研究。
罗大庆, 薛会英, 边巴多吉. 2003. 藏东南色季拉藓类冷杉林群落特征研究[J]. 林业科学研究, 16(增): 7-12. |
李文华. 1985. 西藏森林[M]. 北京: 科学出版社: 38-57.
|
任青山. 2002. 西藏冷杉原始林群落物种多样性初步研究[J]. 生态学杂志, 21(2): 67-70. |
孙玉玲, 李庆梅, 谢宗强. 2005. 濒危植物秦岭冷杉结实特性的研究[J]. 植物生态学报, 29(2): 251-257. DOI:10.17521/cjpe.2005.0032 |
唐润琴, 李先琨, 欧祖兰, 等. 2001. 濒危植物元宝山冷杉结实特性与种子繁殖力初探[J]. 植物研究, 21(3): 403-408. |
谢宗强, 陈伟烈, 胡东, 等. 1998. 濒危植物银杉的结实特性及动物对果实的危害性[J]. 植物生态学报, 22(4): 319-326. |
辛学兵, 翟普明. 2003. 西藏色季拉山冷杉林生态系统的养分循环[J]. 林业科学研究, 16(6): 668-676. |
徐亮, 包维楷, 何永华. 2004. 4个岷江柏种群的球果和种子相态特征及其地理空间差异[J]. 应用与环境生物学报, 10(6): 707-711. |
赵德明. 2001. 中国木本植物种子[M]. 北京: 中国林业出版社: 10-15.
|
Fenner M, Thompson K. 2005. The ecology of seeds[M]. Cambridge: Cambridge University Press.
|
Harper J L. 1977. Population biology of plants[M]. London: Academic Press.
|
Li P, Beaulieu J, Bousquet J. 1997. Genetic structure and patterns of genetic variation among populations in eastern white spruce (Picea glauca)[J]. Canadian Journal of Forest Research, 27(2): 189-198. DOI:10.1139/x96-159 |
Ruby J L. 1967. The correspondence between genetic, morphological and climatic variation pattern in Scotch pine[J]. Silvae Genetica, 16(2): 50-56. |
Steven D D. 1991. Experiments on mechanisms of establishment in old-field succession: seedling emergence[J]. Ecol, 72: 1066-1075. DOI:10.2307/1940606 |
Wheeler N C, Guries R P. 1982. Population structure, genic diversity, and morphological variation in Pinus contorta Dougl[J]. Canadian Journal of Forest Research, 12: 595-606. DOI:10.1139/x82-091 |
Westoby M, Jurado E, Leishman M. 1992. Comparative evolutionary ecology of seed sizes[J]. Trends Ecol Evol, 7: 368-372. DOI:10.1016/0169-5347(92)90006-W |