 2010, Vol. 46
2010, Vol. 46文章信息
- 刘宁, 张芸香, 郭晋平, 张秋枫
- Liu Ning, Zhang Yunxiang, Guo Jinping, Zhang Qiufeng
- 华北落叶松-白杄混交林下更新幼苗幼树的功能特性
- Functional Traits of Naturally Regenerated Seedlings and Saplings of Larix principis-rupprechtii and Picea meyeri in the Mixed Stands
- 林业科学, 2010, 46(7): 22-29.
- Scientia Silvae Sinicae, 2010, 46(7): 22-29.
-
文章历史
- 收稿日期:2009-05-20
- 修回日期:2010-05-24
-
作者相关文章



植物的形态、生理和表型等功能特性影响着植物的个体适应度(fitness),从而间接影响着生态系统的功能(Violle et al., 2007)。群落生态学的发展需要了解这些功能特性如何在异质性环境中控制植物的基本生态位和现实生态位(McGill et al., 2006)。这就要求识别在限制性环境条件下与植物种间表现(performance)差异相关的关键功能特性(Gómez-Aparicio et al., 2007),从而有助于植物物种“功能型”或策略的划分,以了解物种对环境变化的响应和/或物种对生态系统的贡献(Malézieux et al., 2007; Lavorel et al., 2002)。
对于林下更新的树种来说,光照条件是决定其生态位的重要环境因素(Sánchez-Gómez et al., 2006)。木本植物与光环境变化响应相关的功能特性包括高度、地径、比叶面积、地上地下部分生物量、光合特性以及更新特点等(Cornellissen et al., 2003)。这些特性可能决定着林下更新树种在幼龄期表现上的种间差异(Violle et al., 2007)。例如在高光照环境下,阳生树种的净光合速率升高,阴生树种却低于中等光照条件下(Koroleva et al., 2001; Valladares et al., 2002); 在遮荫环境下,阳生树种更倾向于分配更多生物量到地下部分(Landhäusser et al., 2001)、减少对叶组织的生物量分配(Montgomery,2004), 但叶绿素含量和净光合速率保持不变,而阴生树种的叶绿素含量却会下降(Malavasi et al., 2001; 郭晋平等,2009)、比叶面积增大和气孔导度降低(Chen et al., 1996),但净光合速率却可能不低于或高于高光照环境下(Chen et al., 1997)。树种对于光环境在多个不同特性上的响应差异与树种的资源策略有关。一般来说, 阳生树种在形态和光合特性上的较低响应体现出对高光照环境的专一性(Chen et al., 1998),而阴生树种的较大响应表现出对不同光环境的较大适应性(Montgomery, 2004)。因此,研究不同树种的功能特性有助于了解树种对环境的适应能力及其资源利用策略。
幼龄植株对光照响应的种间差异能在很大程度上解释沿演替和生产力梯度的生态位分化(Pacala et al., 1996)。在植物早期的发育中,植物对光照的响应依赖于个体的发育节律(Niinemets, 2006)。研究不同年龄阶段植株的生理生态特性有助于更好地了解环境梯度下树种表现种间差异的机制。
华北落叶松(Larix principis-rupprechtii)和白杄(Picea meyeri)或青杄(Picea wilsonii)的天然混交林是华北山地重要的森林类型,在山西关帝山、管涔山林区阴坡可见保存良好的华北落叶松-白杄(或青杄)混交林(郭晋平等,1997)。一般认为,本地区存在的干扰(林火、砍伐)可能是这2个树种混交林共存的原因。虽然这2个树种林下更新木的更新格局已经确立,但其具体功能特性仍有待研究以全面了解这2个树种的更新机制。本研究旨在调查2种光环境下天然更新华北落叶松和白杄幼苗及幼树的生理和形态响应,通过调查:1)天然更新幼龄华北落叶松和白杄气体交换参数和生长的种间差异; 2)这2个树种在不同光环境下气体交换参数和生长响应的种间差异; 3)这2个树种气体交换参数和生长响应的种间差异是否在幼苗和幼树阶段不同,为森林生态系统的保护和恢复提供科学依据。
1 试验设计和研究方法 1.1 试验地点试验地点选在山西省交城县关帝山林区的庞泉沟自然保护区,位于111°21′—111°37′ E,37°45′—37°59′ N之间,属暖温带落叶阔叶林带的吕梁山北段山地云杉华北落叶松林区。根据1983—1991年气象观测资料(郭晋平等,1997),本地区年均气温4.2 ℃,7月均温17.5 ℃,1月均温-10.2 ℃; 年均降水量822.6 mm; 年均日照时数1 900~2 200 h; 年均蒸发量1 100~1 500 mm。
1.2 试验设计试验采用分裂区组设计,其中主处理为光环境,设置3个重复,次处理为树种和树龄2个因素。试验选择在庞泉沟自然保护区内神尾沟、罗坂沟和西塔沟的华北落叶松-云杉混交林的林窗和林冠下进行,分别代表林窗和林冠下2种光环境。林窗中标准地设在林中空地中,其边界距林缘1倍树高以上,林冠下标准地设在林内,标准地对角线上5个点的光照强度控制在林窗下光强值的40%~60%之间。林窗中(郁闭度约为0)和林冠下(郁闭度约为0.5)分别设置3块标准样地(区组)作为重复,在每块标准地中央,分幼苗和幼树2个树龄,选定华北落叶松和白杄的调查测定样株各3株,共计选定调查测定样株72株。幼苗样株的选定要求符合苗高10~20 cm,苗龄3~4年,幼树样株的选定要求符合树高30~50 cm,苗龄6~8年。
1.3 气体交换参数和生长指标的测定 1.3.1 气体交换参数测定2006年7月初的1周内,在选定植株第2侧枝的当年生、健康、完全展开的成熟叶片上,使用Li-6400系统(LI-COR Inc.,Nebraska,U.S.A)、红蓝光源和标准叶室测定叶片的气体交换参数。测量时,叶室内环境设置为饱和光合有效辐射(saturated PAR)1 200 μmol·m-2s-1、叶温22 ℃、相对湿度60%。所有测量均在10:00—12:00以及14:00—16:00进行。测量后剪下用于气体交换参数测定后的叶片,保存在保温瓶内带回实验室,用WinRhizo分析系统(Regent Instruments Inc.,Quebec City,Quebec,Canada)测定所有叶片的合计投影面积,并根据von Caemmerer等(1981)的公式结合投影面积计算基于叶面积的净光合速率(net photosynthetic rate per unit leaf area, Pn area)、气孔导度(gs)、叶片蒸腾速率(E)及胞间/大气CO2浓度比(Ci/Ca)。基于叶干质量的净光合速率(net photosynthetic rate per unit leaf mass, Pn mass)为Pn area和比叶面积(SLA)的乘积。即时光合水分利用效率(WUE)为基于叶面积的净光合速率与叶片蒸腾速率之比。
1.3.2 生长指标的测定在完成气体交换参数指标的测定后,逐一测量所选样株的高度和地径,剪取植株的茎部和叶片带回实验室,在80 ℃下烘干24 h,用感量0.000 1 g的电子天平测定茎干质量(SDW)和叶片干质量。比叶面积(SLA)为投影面积和叶片干质量之比。
1.3.3 数据分析根据试验设计,为检验光照条件、树种和年龄阶段对测定指标的影响和交互作用,使用以下线性模型分别对测得的气体交换参数和生长指标进行三因素方差分析:
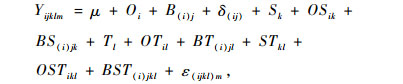
|
式中,Yijklm是特定应变量的测量值; μ是应变量的总平均值; Oi是2种光照处理之一的固定效应; B(i)j是区组的随机效应; δ(ij)是全区组限制误差; Sk是2个树种之一的固定效应; OSikl是光照条件和树种的交互作用; BS(i)jk是区组和树种的交互作用; Tl是2个树龄之一的固定效应; OTil是光照条件和树龄的交互作用; BT(i)jl是区组和树龄的交互作用; STkl是树种和树龄的交互作用; OSTikl是光照条件、树种和树龄三者的交互作用; BST(i)jkl是区组、树种和树龄三者的交互作用; ε(ijkl)m是抽样误差。
为使数据符合正态分布,满足统计检验要求,用自然对数变换y′= ln(y+1)对地径和株高数据进行预处理,处理后的数据用SAS/STAT 8.0分析软件(SAS Institute Inc, Cary, NC, USA)中的PROC GLM程序、RANDOM/TEST选项进行显著性检验,并对差异显著(P<0.05)的指标用LSD检验进行多重比较。
2 结果 2.1 不同光照条件下华北落叶松和白杄幼苗及幼树气体交换参数间的差异对不同光照条件下华北落叶松和白杄幼苗及幼树的气体交换参数进行方差分析(表 1)。
|
|
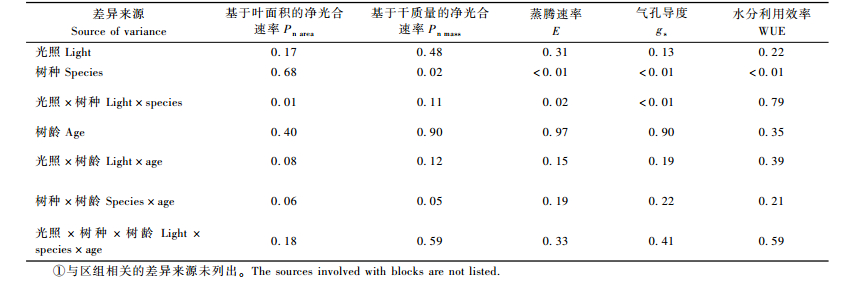
由表 1可见,不同光照或树龄之间,样株的气体交换参数均无显著差异(P>0.05),而在树种之间,华北落叶松和白杄的气体交换参数之间的差异极显著(P<0.01,基于叶面积的净光合速率除外)。
由表 2可见,在选择的试验区内,华北落叶松基于叶面积的净光合速率平均值与白杄相似,而其基于叶干质量的净光合速率比白杄低37%,平均水分利用效率(WUE)低36%,而华北落叶松的平均气孔导度(gs)和蒸腾速率(E)却分别比白杄高55%和47%。这一结果说明白杄的净光合速率并不低于华北落叶松,且在水分利用方面优于华北落叶松。
|
|

在不同的光照条件下,华北落叶松和白杄基于叶面积的净光合速率、气孔导度和蒸腾速率显著不同(图 1,P<0.05)。随着光照的减弱,华北落叶松基于叶面积的净光合速率显著升高(74%),而白杄却在这2种光照条件下保持一致。另外,在林窗下华北落叶松基于叶面积的净光合速率显著低于白杄(22%),而在林冠下却显著高于白杄(20%)。同时,华北落叶松的气孔导度和蒸腾速率均随光照减弱显著升高(分别升高106%和59%),而白杄的这2个参数却始终保持一致,与林窗下的华北落叶松相同。

|
图 1 林窗和林冠下华北落叶松和白杄基于叶面积的净光合速率、气孔导度及蒸腾速率 Figure 1 Pn area, gs and E of L. principis-rupprechtii and P. meyeri growing under open and understory light regimes |
华北落叶松和白杄幼苗及幼树基于叶干质量的净光合速率均无显著差异,但华北落叶松表现出幼树略低于幼苗的趋势,白杄则是幼树略高于幼苗(图 2)。

|
图 2 华北落叶松和白杄幼苗及幼树基于叶干质量的净光合速率 Figure 2 Pn mass of L. principis-rupprechtii and P. meyeri seedlings and saplings |
对不同光照条件下华北落叶松和白杄幼苗及幼树的地径、高度、比叶面积、高径比和茎干质量进行方差分析(表 3)。由表 3可见,光照条件对地径、高度、比叶面积、高径比和茎干质量等生长特征均无显著影响(P>0.05)。除树高外,华北落叶松和白杄的其他生长特征存在显著或极显著差异(P<0.05或P<0.01)。
|
|
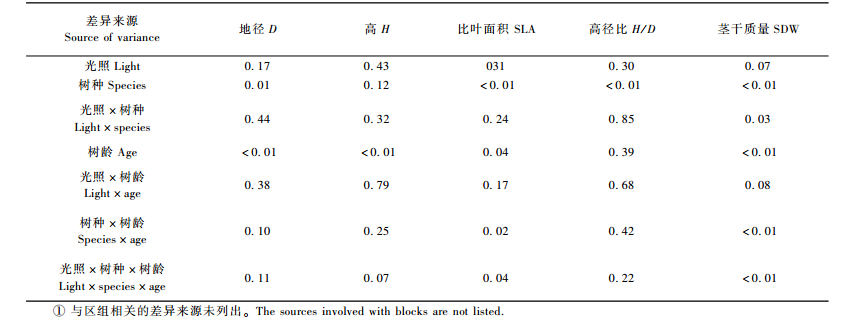
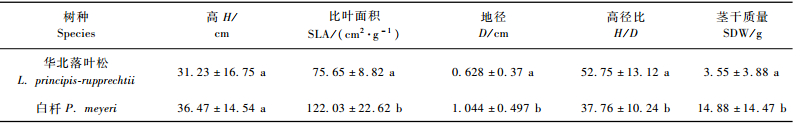
由表 4可见,幼龄(3~8年)华北落叶松的平均地径和茎干质量分别比白杄低40%和76%,而高径比却高出白杄40%。华北落叶松的比叶面积也比白杄低39%。这一结果表明白杄在生长指标上优于华北落叶松。
|
|
对茎干质量来说,光照和树种的交互作用、树种和树龄的交互作用以及光照、树种和树龄3者的交互作用均显著或极显著(分别为P<0.05, P<0.01, P<0.01,表 2)。另外在比叶面积指标上,树种和树龄的交互作用以及光照、树种和树龄3者的交互作用也显著(P<0.05)。
由图 3可见,林窗和林冠下华北落叶松的茎干质量并无显著差异(P>0.05),而白杄在林窗下的茎干质量几乎是林冠下的2倍(P<0.05)。由图 3可见,单从树龄角度看,3~4年生幼苗的茎干质量在林窗和林冠下并无显著区别(P>0.05),而对于6~8年生的幼树来说,林窗下的茎干质量却是林冠下的2倍(P<0.05)。

|
图 3 林窗和林冠下华北落叶松和白杄及其幼苗和幼树的茎干质量 Figure 3 Stem dry weights (SDW) of L. principis-rupprechtii and P. meyeri and the combined SDW of seedlings and saplings growing under open and understory light regimes |
由图 4可见,华北落叶松幼苗和幼树的比叶面积始终低于白杄。随着白杄的生长,幼树的比叶面积高出幼苗12%左右。

|
图 4 华北落叶松和白杄幼苗和幼树的比叶面积 Figure 4 Specific leaf area (SLA) of L. principis-rupprechtii and P. meyeri seedlings and saplings |
由图 5可见,华北落叶松和白杄幼苗的茎干质量在林窗和林冠下分别无显著差异(P>0.05)。另外,华北落叶松幼树的茎干质量与幼苗相比在林窗和林冠下分别增加5.64 g和5.13 g,并无显著差异,而白杄幼树的茎干质量与幼苗相比,在林窗下和林冠下分别增加28.51 g和11.26 g,前者是后者的2.5倍。同时,白杄和华北落叶松幼树茎干质量之间的差距从林窗下的27.6 g下降到林冠下的9.57 g,降低约65%。由图 5可见,华北落叶松和白杄幼苗及幼树的比叶面积在林窗和林冠下无变化。白杄幼树的比叶面积在林窗环境下高出幼苗19%,而在林冠下两者无差异。

|
图 5 光照、树种和树龄三者交互作用下华北落叶松和白杄幼苗及幼树的茎干质量和比叶面积 Figure 5 SDW and SLA of L. principis-rupprechtii and P. meyeri seedlings and saplings growing under open and understory light regimes |
阳生树种和阴生树种在气体交换参数和生长特征上存在明显差异,例如阳生树种叶片的暗呼吸速率更高、净光合速率更大、比叶面积更小、高径比更大等(Chen et al., 1997; 1998; Guo et al., 2004)。关于幼龄木本植株的研究表明,幼苗幼树的耐荫性与以牺牲生长潜力而换取长期生存的保守资源利用策略有关,即阴生树种可表现出低比叶面积、低叶面积比、低高径比、低暗呼吸速率和高根茎生物量分配的特性,而不一定是与高碳同化效率相关的特性(Sánchez-Gómez et al., 2006; 2007; Reich et al., 1998; 2003)。本研究中,除了基于叶面积的净光合速率相同外,林下天然更新的华北落叶松和白杄在气体交换参数方面表现出明显的种间差异。阴生树种白杄的蒸腾速率、气孔导度、高径比低于华北落叶松,水分利用效率和茎生物量则高于华北落叶松,与上述保守资源利用策略相一致,体现出一定的耐荫性。另一方面,白杄基于叶干质量的净光合速率高于华北落叶松,同时比叶面积也相对较大,表现出与高碳同化能力相关的特性,与保守的资源利用策略不一致。这种现象曾报道于几个阴生的栎树(Quercus)和阳生的松树(Pinus)树种中,可能与树种特定的叶结构特征(Sánchez-Gómez et al., 2006)和叶绿素含量(Hansen et al., 2002)相关。
3.2 光照处理下树种响应的种间差异光照条件是决定树种更新生态位的重要环境因素(Gómez-Aparicio et al., 2006)。不同光照条件下光合效率和光合产物分配模式的差异在很大程度上影响不同树种的幼年期生长(Montgomery,2004),另外也有可能影响树种的更新模式(Grassi et al., 2001)。一般来说,随着光照强度的增加,阳生树种的光合效率升高,阴生树种的光合效率反而会低于中等光照条件下(Koroleva et al., 2001; Valladares et al., 2002; 郭晓荣等,2004)。本研究中,华北落叶松在林窗环境下的净光合速率显著低于白杄,在林冠下则显著升高并高于白杄,同时2个树种基于叶干质量的净光合速率的响应却无变化,并未表现出一致的趋势。对于天然环境下的净光合速率来说,除光照条件外,林冠下微环境中的水分条件、土壤等其他环境因素又常常与光照产生复杂的交互作用(Chen et al., 1997)。同时,本研究中华北落叶松的蒸腾速率和气孔导度在林冠条件下分别升高59%和106%,均与其净光合速率紧密相关(r2分别为0.85和0.76,P<0.01)。由此可见,林窗环境下华北落叶松较低的净光合速率可能与不利的微环境水分条件密切相关。另外,华北落叶松虽属速生树种,但在苗期需要一定的遮荫才能生长良好(郭晋平等,2009),而且阳生树种苗期遮荫条件下生长情况好于全光照条件下的现象也可在同属落叶松属的西部落叶松(Larix occidentalis)野外人工栽植幼苗中见到(Chen et al., 1998; Schmidt et al., 1990)。白杄基于叶面积和基于叶干质量的净光合速率在林窗和林冠环境下均相同,除非受到光抑制, 同阴生树种栽植于遮荫环境下与林窗环境下并无劣势的结论相一致(Chen et al., 1997)。
低光照环境一般导致木本植物的生物量积累降低,并分配更多生物量到地下部分(Groninger et al., 1996; Wiebel et al., 1994),并且减少对叶组织的生物量分配(Montgomery,2004),但也有例外情况,如北美鹅掌楸(Liriodendron tulipifera)能在低光照下获得更高的茎生长(Groninger et al., 1996)。也有研究认为幼苗茎生物量的投入与光照关系不密切,而与树种本身有关(Poorter, 1999)。本研究中华北落叶松的生长指标在林冠环境下未受明显抑制,未对光照处理产生响应,而林冠下白杄的茎干质量下降到林窗下的一半左右,对光照产生较大的形态响应,缩小了华北落叶松和白杄之间的形态差距。
华北落叶松的气体交换参数在2种光环境下表现出较大的响应差异,在生长指标上响应差异较小,符合生理响应差异与较高的占据林窗能力和在高光照环境下生长能力相关的结论,而白杄在气体交换参数上响应差异较小,在生长指标上较大,符合形态响应差异与林下生存和生长能力相关的结论(Valladares et al., 2002; Niinemets et al., 2004)。另外,华北落叶松和白杄的生理形态特性与以往研究中不同试验条件光环境下分别单株栽植的存活率相对应(郭晋平等,2009)。然而,本研究未调查华北落叶松和白杄的生物量和生物量分配指标,另外野外研究中也很难控制水分、土壤和营养等协变量的影响,因此此生理生态的响应差异结论需要通过进一步研究确认。
3.3 树龄对光照处理下树种响应种间差异的影响随着树龄的增加(从3~4年到6~8年),华北落叶松基于叶干质量的净光合速率呈下降趋势,白杄略有上升,但并未达到显著水平。在茎干质量指标上,这2个树种对光照产生截然不同的响应。对于华北落叶松来说,2种光照条件下的茎干质量并无不同,且在6~8年时达到的茎干质量也相同; 对白杄来说,虽然3~4年时的茎干质量在2种光照条件下并无区别,但在6~8年时,林窗条件下的茎干质量增长却高出林冠下增长1.5倍左右,表明林冠条件明显抑制了白杄幼树的茎生长。同时,华北落叶松的比叶面积未对光照和树龄产生任何响应,而白杄幼树的比叶面积仅在林窗环境下较幼苗显著增大,同样的结果可见于阴生的夏栎(Quercus robur)中(Sánchez-Gómez et al., 2006)。华北落叶松和白杄幼苗和幼树的光响应差异说明树种在生长发育中对限制性环境因子的响应可能会发生变化,从而影响两者的共存机制。尽管如此,植物早期发育中对光照的响应依赖于个体的发育,早期确立的关系和限制因素可能在随后的生长阶段中变化,并导致树种相对生长速率的变化(Niinemets, 2006)。
3.4 结论1) 林下天然更新的幼龄华北落叶松和白杄在气体交换参数和生长特征上表现出显著差异。作为阴生树种的白杄在光合效率、水分利用和生长上均优于华北落叶松,并未表现出专一的保守资源利用策略。
2) 华北落叶松和白杄在对不同光环境的生理和形态响应上分别表现出较大的差异。华北落叶松在气体交换参数上产生对光照条件的显著响应,但在生长特征上无响应。白杄的气体交换参数并未受到光环境变化的影响,而生长则在遮荫环境下受到显著抑制。因水分、土壤和营养等协同因素的存在,此结论仍需进一步研究确认。
3) 随着树龄的增加,林冠下白杄幼树的茎干质量和比叶面积低于林窗环境下,表现出随生长而发生变化。
郭晋平, 王石会, 康日兰. 1997. 管涔山青杄(Picea wilsonii)天然林年龄结构及其动态的研究[J]. 生态学报, 17(2): 184-189. |
郭晋平, 李海波, 刘宁, 等. 2009. 华北落叶松(Larix principis-rupprechtii)和白杄(Picea meyeri)幼苗对光照和竞争响应的差异比较研究[J]. 林业科学, 45(2): 53-59. DOI:10.11707/j.1001-7488.20090209 |
Canham C D. 1988. Growth and canopy architecture of shade-tolerant trees: response to canopy gaps[J]. Ecology, 69: 786-795. DOI:10.2307/1941027 |
Chazdon R L. 1986. Light variation and carbon gain in rain forest understory palms[J]. J Ecol, 74: 995-1012. DOI:10.2307/2260229 |
Chazdon R L. 1988. Sunflecks and their importance to forest understory plants[J]. Adv Ecol Res, 18: 1-63. DOI:10.1016/S0065-2504(08)60179-8 |
Chen H Y, Klinka K, Kayahara G J. 1996. Effects of light on growth, crown architecture and specific leaf area for naturally established Pinus concorta var. latifolia and pseudotsuga menziesii var. glauca saplings[J]. Can J For Res, 26: 1149-1157. DOI:10.1139/x26-128 |
Chen H Y, Klinka K. 1997. Light availability and photosynthesis of pseudotsuga menziesii seedlings grown in the open and in the forest understory[J]. Tree Physiology, 17: 23-29. DOI:10.1093/treephys/17.1.23 |
Chen H Y, Klinka K. 1998. Survival, growth and allometry of planted Larix occidentalis seedlings in relation to light availability[J]. For Eco Mang, 106: 169-179. DOI:10.1016/S0378-1127(97)00309-5 |
Cornellissen J H C, Lavorel S, Garnier E, et al. 2003. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide[J]. Australian Journal of Botany, 51: 334-377. |
Gómez-Aparicio L, Valladares F, Zamora R. 2006. Differential light responses of Mediterranean tree saplings: linking ecophysiology with regeneration niche in four co-occurring species[J]. Tree Physiology, 26: 947-958. DOI:10.1093/treephys/26.7.947 |
Groninger J W, Seiler J R, Peterson J A, et al. 1996. Growth and photosynthetic responses of four Virginia Piedmont tree species to shade[J]. Tree Physiology, 16: 773-778. DOI:10.1093/treephys/16.9.773 |
Grassi G, Bagnaresi U. 2001. Foliar morphological and physiological plasticity in Picea abies and Abies alba saplings along a natural light gradient[J]. Tree Physiology, 21: 959-967. DOI:10.1093/treephys/21.12-13.959 |
Guo X R, Cao K F, Xu Z F. 2004. Response of photosynthesis and antioxygenic enzymes in seedlings of three tropical forest tree species to different light environment[J]. Chin J Appl Ecol, 15(3): 377-381. |
Hansen U, Schneiderheinze J, Rank B. 2002. Is the lower shade tolerance of scots pine, relative to pedunculate oak, related to the composition of photosynthetic pigments?[J]. Photosynthetica, 40: 369-374. DOI:10.1023/A:1022670807608 |
Koroleva K G H, Dalling O Y, Winter J W. 2001. Acclimation of tropical tree seedlings to excessive light in simulated tree-fall gaps[J]. Plant Cell Environ, 24(12): 1345-1352. DOI:10.1046/j.0016-8025.2001.00786.x |
Landhäusser S M, Lieffers V J. 2001. Photosynthesis and carbon allocation of six boreal tree species grown in understory and open conditions[J]. Tree Physiology, 21: 243-250. DOI:10.1093/treephys/21.4.243 |
Lambers H, Chapin Ⅲ F S, Pons T L. 1998. Plant physiological ecology[M]. New York: Springer-Verlag New York Inc.
|
Lavorel S, Garnier E. 2002. Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail[J]. Functional Ecology, 16: 545-556. DOI:10.1046/j.1365-2435.2002.00664.x |
Malavasi U C, Malavasi M M. 2001. Leaf characteristics and chlorophyll concentration of Schyzolobium parahybum and Hymenaea stilbocarpa seedlings grown in different light regimes[J]. Tree Physiology, 21: 701-703. DOI:10.1093/treephys/21.10.701 |
Malézieux E, Lamanda N, Laurans M, et al. 2007. Plant functional traits and types: their relevance for better understanding the functioning and properties of agroforestry systems. Second Inter-national Symposium on Multi-Strata Agroforestry Systems with Perennial Crops. Turrialba, Costa Rica. https://www.sciencedirect.com/science/article/pii/S1360138517300195
|
McGill B J, Enquist B J, Weiher E, et al. 2006. Rebuilding community ecology from functional traits[J]. Trends in Ecology and Evolution, 21: 178-185. DOI:10.1016/j.tree.2006.02.002 |
Messier C, Puttonen P. 1995. Growth, allocation and morphological responses of Betula pubescens and Betula pendula to shade in developing scots pine stands[J]. Can J For Res, 25: 629-637. DOI:10.1139/x95-070 |
Montgomery M. 2004. Relative importance of photosynthetic physiology and biomass allocation for tree seedling growth across a broad light gradient[J]. Tree Physiology, 24: 155-167. DOI:10.1093/treephys/24.2.155 |
Niinemets Ü. 2006. The controversy over traits conferring shade tolerance in trees: entogenetic changes revisited[J]. J Ecol, 94: 464-470. DOI:10.1111/jec.2006.94.issue-2 |
Niinemets Ü, Valladares F. 2004. Photosynthetic acclimation to simultaneous and interacting environmental stresses along natural light gradients: optimality and constraints[J]. Plant Biol, 6: 254-268. DOI:10.1055/s-2004-817881 |
Pacala S W, Canham C D, Saponara J, et al. Forest models defined by field measurements: estimation, error analysis, and dynamics[J]. Ecol Monogr, 66: 1-43. DOI:10.2307/2963479 |
Poorter L. 1999. Growth response of 15 rainforest tree species to a light gradient: The relative importance of morphological and physiological traits[J]. Funct Ecol, 13: 396-410. DOI:10.1046/j.1365-2435.1999.00332.x |
Sánchez-Gómez D, Zavala M A, Valladares F. 2006. Seedling survival responses to irradiance are differentially influenced by drought in four tree species of the Iberian cool temperate-Mediterranean ecotone[J]. Acta Oecol, 30: 322-332. DOI:10.1016/j.actao.2006.05.005 |
Sánchez-Gómez D, Zavala M A, Valladares F. 2007. Functional traits and plasticity linked to seedlings'performance under shade and drought in Mediterranean woody species[J]. Ann For Sci, 65: 311-321. |
Schmidt W C, Shearer R C. 1990. Larix occidentalis Nutt.-Western larch//Burns R M, Honkala H B. Silvics of North America: Vol. 1. Conifers. Agricultural Handbook. Washington DC: USDA, Forest Service, 160-172.
|
Valladares F, Wright S J, Lasso E, et al. 2000. Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest[J]. Ecology, 81: 1925-1936. DOI:10.1890/0012-9658(2000)081[1925:PPRTLO]2.0.CO;2 |
Valladares F, José M C, Ismael A. 2002. The greater seedling highlight tolerance of Quercus robur over Fagus sylvatica is linked to a greater physiological plasticity[J]. Trees, 16: 395-403. |
Violle C, Navas M L, Vile D, et al. 2007. Let the concept of trait be functional![J]. Oikos, 116: 882-892. DOI:10.1111/oik.2007.116.issue-5 |
von Caemmerer S, Farquhar G D. 1981. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves[J]. Planta, 153: 376-387. DOI:10.1007/BF00384257 |
Wiebel J, Chacko E K, Downton W J S, et al. 1994. Influence of irradiance on photosynthesis, morphology and growth of mangosteen (Garcinia mangostana L.) seedlings[J]. Tree Physiology, 14: 263-274. DOI:10.1093/treephys/14.3.263 |