 2010, Vol. 46
2010, Vol. 46文章信息
- 吴甘霖, 王志高, 段仁燕, 张中信, 沈三保
- Wu Ganlin, Wang Zhigao, Duan Renyan, Zhang Zhongxin, Shen Sanbao
- 安徽大别山多枝尖山区植物物种多样性
- Species Diversity of Plant Community in Duozhijian Region of Dabieshan Mountain Anhui Province
- 林业科学, 2010, 46(6): 128-132.
- Scientia Silvae Sinicae, 2010, 46(6): 128-132.
-
文章历史
- 收稿日期:2009-12-31
- 修回日期:2010-04-29
-
作者相关文章


2. 鹞落坪国家级自然保护区管委会 安庆 246600
2. Yaoluoping Reserve Administration Committee Anqing 246600
群落物种多样性反映了生物群落在组成、结构、功能和动态方面的异质性,是生态学研究中十分重要的内容(兰思仁,2003;Ricklefs,2004;井学辉等,2010)。大别山位于安徽、湖北和河南3省交界处,是长江和淮河的天然分水岭,其地带性植被属由亚热带常绿阔叶林向暖温带落叶阔叶林过渡类型(谢中稳等,1995)。多枝尖山区是大别山腹地主峰分水岭的主段,是大别山的第三高峰,最高海拔1 721.5 m。该区地带性植被为常绿落叶阔叶混交林,植物区系起源古老,属华中、华西、华北、华南和华东植物区系的交汇地带,特有现象明显,是大别山植物区系物种最丰富的地区之一,也是南北植物迁徙的主要通道之一(刘宾,1991;单人驊等,1964;谢中稳等,1994;1995;高林等,2002)。有关这一区域的植被和植物区系曾有过报道(谢中稳等,1993;谢中稳,1994;高林等,2002;高林,2004)。但缺乏对群落结构和物种多样性的研究。
本研究在对多枝尖山区植植物群落充分调查的基础上,采用TWINSPAN法对该区植被进行群落类型分类,分析各群落类型的群落结构和物种多样性,以此为依据进一步分析该区植被恢复中的演替规律,为植被保护提供科学依据。
1 研究区概况多枝尖山区位于安徽省岳西县境内,西部与湖北省英山县交界,为大别山腹地主峰分水岭的主段(116°03′—116°09′E,31°00′—31°11′N),总面积约210 km2。多枝尖山区在20世纪70年代曾遭到大规模砍伐,目前是1994年设置的鹞落坪国家级自然保护区的核心区之一,次生植被恢复良好。该区属于华东亚热带气候湿润气候区,年平均气温12.7 ℃,年降水量1 600 mm,全年无霜期约195天;山体主要由花岗岩组成,并夹有少量花岗岩和角闪片麻岩;在海拔800 m以上以山地棕壤为主,海拔800 m以下主要为山地黄棕壤,一般呈中性或微酸性,局部地区有草甸土和沼泽土分布。据谢中稳等(1993)报道,该区在海拔800 m以下分布有常绿、落叶阔叶混交林,常绿树种有青冈栎(Cyclobalanopsis glauca)、冬青(Ilex purpurea)和豺皮樟(Litsea rotundifolia)等,落叶树种有短柄枹(Quercus serrata)、木蜡树(Toxicodenddron sylvestre)和扁担杆(Grewia biloba)等;在海拔800~1500 m分布着落叶阔叶林带,群落优势种有江南桤木(Alnus trabeculosa)、茅栗(Castanea seguinii)和华千金榆(Carpinus cordata)等;黄山松(Pinus taiwanensis)分布于海拔800 m以上,常形成纯林;区系成分丰富,特有现象明显,以温带为主,表现出由温带向亚热带过渡的特征。
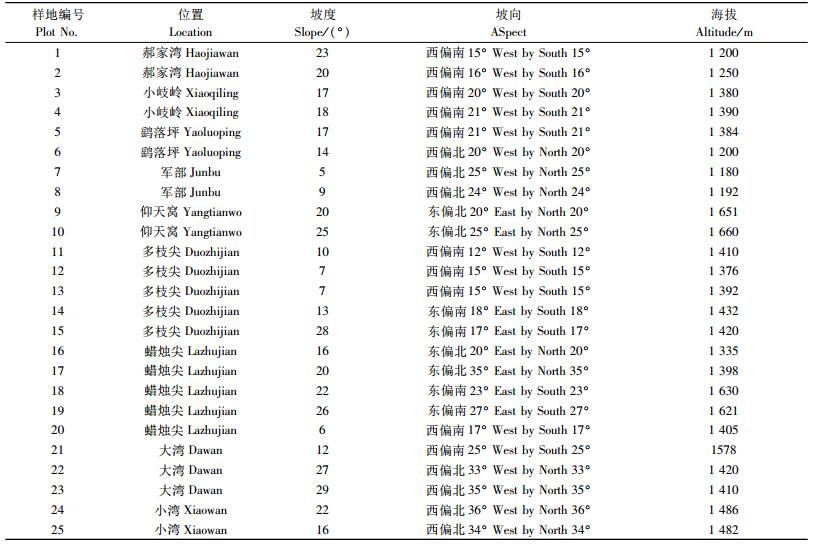
2 研究方法在研究区域典型地段共设置25个20 m×20 m的乔木样地,样地概况见表 1。每个乔木样地中设置5个2 m×2 m的灌木样方和5个1 m×1 m草本样方。对乔木层逐株调查,记录种名、胸径和坐标;对灌木层和草本层记录种名、株数、高度和盖度,同时记录各样地的坡度、坡向和海拔等环境数据。群落类型的划分采用TWINSPAN法。
|
|
本研究以重要值(乔木层重要值为相对密度、相对频度和相对显著度3者之和;灌木层及草本层重要值为相对密度、相对频度和相对盖度3者之和)作为多样性指数的测度依据,选用Shannon-Wiener多样性指数(H)、Simpson多样性指数(C)和Pielou均匀度指数(Jsw)作为多样性指数:
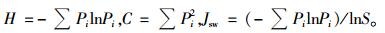
|
式中:Pi为物种i的相对重要值,S为物种i所在样地的物种总数目。
3 结果与分析 3.1 群落类型的划分及其特征根据TWINS-PAMN数量分类方法将25个乔木样地划分为9个群落类型。TWINSPAN分类结果树状图见图 1,各群落类型环境和物种组成情况见表 2。

|
图 1 TWINSPAN分类树状图 Figure 1 Dendrogram of TWINSPAN |
|
|

1) 华山松(Pinus armandii)+映山红(Rhododendron simsii)+苔草(Carex sp.)群落 该群落类型位于郝家湾,包括样地1和2,海拔1 250 m,位于阳坡。乔木层共记录到23个物种,除华山松优势度较大外,还有黄山松、杉木(Cunninghamia lanceolata)、茅栗和短柄枹等。灌木层种类较多,共有33个物种,以映山红占优势,另外有山胡椒(Lindera glauca)、杜鹃(Rhododendron daurici)和山茶(Camellia japonica)等,该层中短柄枹幼树较多。草本层以苔草占优势,另有野青茅(Deyeuxia arundinacea)、求米草(Oplismenus undulatifolius)和中华委陵菜(Potentilla chinensis)等。
2) 华山松+山莓(Rubus corchorifolius)+求米草群落 这一群落类型见于小岐岭以及鹞落坪自然保护区管理处周围,包括样地2, 4和5。该群落类型的乔木层共有35个物种,华山松为20世纪80年代人工种植,常见伴生种有白檀(Symplocos baniculata)、黄山松和化香(Platycarya strobilacea)等。灌木层以山莓占绝对优势,常见种还有山樱花(Cerasus serrulata)、山胡椒以及较多的白檀幼树。草本层常见植物有金星蕨(Parathelypteris glanduligera)、堇菜(Viola verecunda)和鳞毛蕨(Kuniwatsukia sp.)等。
3) 金钱松(Pseudolarix amabilis)+山胡椒(Lindera glauca)+金星蕨群落 该金钱松林为人工种植,包括样地6,7和8。海拔1 180 m,坡向西偏北,坡度5~14°。常见乔木层伴生种有茅栗、黄山松和山樱花等;灌木层以山胡椒为优势种,伴生种有山莓(Rubus corchorifolius)、山橿(Lindera reflexa)和白檀幼树等;草本层常见伴生种有堇菜、鳞毛蕨和求米草等。
4) 黄山栎(Quercus stewardii)+箬竹(Indocal-amus latifolius)+冷水花(Pilea notata)群落 该群落类型位于仰天窝,包括样地9和10,海拔1 651~1 660 m,人迹罕至。乔木层以黄山栎占优势,常见物种还有水马桑(Wigela japonica)、白檀、华东椴(Tilia japonica)、安徽槭(Acer anhweiense)、四照花(Cronus japonica)和省枯油(Staphyleaceae sp.)等;灌木层以箬竹占绝对优势,其次为凤尾竹(Bambusa glancescens)、山胡椒以及省枯油和白檀幼树;草本层优势种为冷水花,常见伴生种有牯岭凤仙(Impatiens davidi)、鳞毛蕨和一把伞天南星(Arisaema erubescens)等。
5) 黄山松+山莓+苔草群落 该群落分布于多枝尖,包括样地11,12和13。海拔1 370~1 410 m。乔木层以黄山松为优势种,伴生种有金钱松、白檀、鹅耳枥(Carpinus turczaninowii)、茅栗(Castanea seguinii)和灯台树(Cornus controversa)等;灌木层以山莓为优势种,伴生种还有三桠乌药(Lindera obtusiloba)、粉花绣线菊(Spiraea japonica)和山胡椒等。草本层以苔草为优势种,伴生种有芒草(Miscanthus sp.)、金星蕨、林荫千里光(Senecio nemorensis)和鳞毛蕨等。
6) 短柄枹+映山红+苔草群落 该群落类型多位于山脊上,包括样地16和17。乔木层以短柄枹占优势,还分布有茅栗、灯台树和四照花(Dendronenthamia japonica)等;灌木层以映山红为优势种,常见物种还有山橿和五味子(Schisandra chinesis);草本层以苔草为优势种,常见种还有芒草、鳞毛蕨和萱草(Hemerocallis fulva)等。
7) 黄山松+三桠乌药+芒草群落 该群落类型包括样地20和21。乔木层以黄山松占绝对优势,其次为白檀和白栎(Quercus fabri)。灌木层除三桠乌药占优势外,常见种有映山红和茶荚蒾(Viburnum setigerum)等,另外白檀、茅栗、漆树(Toxicodendron sylvestris)、四照花和化香等乔木的幼树也较多;草本层以芒草占绝对优势,常见的伴生种有林荫千里光、白穗花(Speirantha gardenii)和鳞毛蕨等。
8) 短柄枹+映山红+芒草群落 该群落类型包括样地14,15,18和19,分布于海拔1 420~1 630 m。乔木层以短柄枹为优势种,常见种有野鸦椿(Euscaphis japonica)、漆树和垂珠花(Styrax dasyanthus)等。灌木层除映山红占优势外,常见种有山樱花、三桠乌药和五味子,以及较多的漆树幼苗。草本层种类较少,除了苔草占优势外,堇菜和禾草(Poa sp.)也较常见。
9) 黄山松+映山红+苔草 该群落类型包括样地22,23,24和25。分布于海拔1 410~1 486 m。乔木层以黄山松为优势种,常见种还有短柄枹、白檀和四照花;灌木层以映山红占绝对优势,常见物种还有山樱花、山梅和三桠乌药等,另外白檀、漆树和短柄枹的幼树也较多。草本层除苔草占优势外,常见物种还有芒草、一枝黄花(Solidago decurens)和珍珠菜(Lysimachia clethroides)等。
3.2 物种多样性根据上述群落类型的划分结果,分别计算出各群落类型中乔木层、灌木层和草本层的物种多样性指数(表 3)。Simpson指数对常见种较为敏感,而Shannon-Wiener指数对稀少种敏感(谢晋阳等,1994)。短柄枹+映山红+苔草群落中乔木层的Shannon-Wiener指数大于3.5。一般认为Shannon-Wiener指数为1~3.5比较准确。本研究区域内植被次生现象比较明显,随着植被的恢复,阔叶树种逐渐侵入针叶林中,且该区位于亚热带北缘,分布有较多的特有物种(谢中稳等,1993),这些因素可能导致了多枝尖山区植物群落中稀少种较多,因此对该区域群落物种多样性进行分析时,Shannon-Wiener指数不适用。
|
|
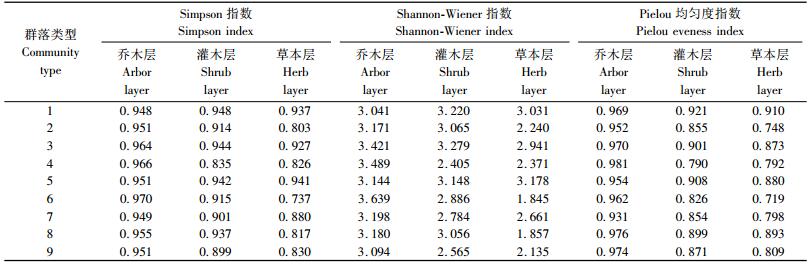
对9种群落类型物种多样性指数的分析表明,乔木层Simpson指数和Pielou均匀度指数均最高,灌木层次之,草本层最低(表 2和3)。而物种丰富度则趋势不明显,华山松+山莓+求米草群落、金钱松+山胡椒+金星蕨群落、短柄枹+映山红+苔草群落和短柄枹+映山红+芒草群落中均是灌木层物种丰富度最高,3种阔叶林群落(短柄枹+映山红+芒草群落、黄山栎+箬竹+冷水花群落和短柄枹+映山红+芒草群落)中草本层的物种丰富度均最低。
物种丰富度指数还表明,乔木层中除短柄枹+映山红+芒草群落外,阔叶林群落的物种丰富度高于针叶林,同是针叶林的金钱松林(金钱松+山胡椒+金星蕨)高于黄山松林(黄山松+山莓+苔草群落、黄山松+三桠乌药+芒草群落和黄山松+映山红+苔草群落)和华山松林(华山松+映山红+苔草群落和华山松+山莓+求米草群落);灌木层中金钱松林物种丰富度最高,华山松林次之,2者均高于黄山松林和阔叶林类型群落;草本层中短柄枹林(短柄枹+映山红+芒草群落群落和短柄枹+映山红+芒草群落)的物种丰富度最低,除了黄山松+映山红+苔草群落外,针叶林群落物种丰富度高于阔叶林群落(表 3)。
4 结论与讨论TWINSPAN分类结果表明,多枝尖山区植物群落样地可分为9个类型。不同类型的群落中群落结构和物种多样性存在差异,而这种差异与各类型群落所处的环境、所处的演替阶段以及人为干扰程度有关(高林等,2002;高林,2004;徐慧等,2002;Getzin et al., 2008)。植物群落上层树种分布的空间异质性会影响到幼苗生长和林下植物的组成(Maltez-Mouro et al., 2005),在植物群落从针叶林、针阔混交林到阔叶林演替的不同阶段,其物种多样性指数依次升高(兰思仁,2003),但演替最高阶段的物种多样性并不是最高(谢晋阳,1994;黄忠良等,2000)。
通常,亚热带森林生态系统群落中林下木本植物物种多样性明显高于乔木层(彭少麟等,1983;吴承祯等,1996)。本研究表明所有群落类型中都是乔木层的Simpson指数和Pielou均匀度指数最高,草本层最低,这一现象很可能是本研究区域植被的次生特性的反映。本研究还表明在研究区域内阔叶林下的草本层物种丰富度最低,这一方面与乔木层和灌木层的物种多样性高有关(Klinka et al., 1996;Maltez-Mouro et al., 2005;Messier,1998),另一方面也与阔叶林下某些物种的优势度过大有关(黄山栎林下灌木层中箬竹占的优势度非常大,使得该群落草本层植物稀少,只有20个物种;短柄枹林下映山红数量较多,使得其草本层植物种类较少)。
本研究结果对深入了解多枝尖山区乃至大别山区植被分布具有重要意义,并为该区植被保护提供科学依据。今后还应进一步探讨影响该区植物群落物种多样性的因素(如土壤、地形和人为干扰等)及该区植被演替规律。
高林. 2004. 皖西南鹞落坪国家级自然保护区特有植物的调查研究[J]. 安徽农业科学, 32(6): 1184-1186. |
高林, 徐如松, 曹姣仙, 等. 2002. 鹞落坪国家级自然保护区植被类型的初步探讨[J]. 生物学杂志, 19(6): 32-42. |
黄忠良, 孔国辉, 何道泉. 2000. 鼎湖山植物群落多样性的研究[J]. 生态学报, 20(2): 193-198. |
井学辉, 臧润国, 丁易, 等. 2010. 新疆阿尔泰山小东沟北坡植物多样性沿海拔梯度分布格局[J]. 林业科学, 46(1): 23-28. DOI:10.11707/j.1001-7488.20100104 |
兰思仁. 2003. 武夷山国家级自然保护区植物物种多样性研究[J]. 林业科学, 39(1): 36-43. |
刘宾. 1991. 安徽大别山陀尖山区植物区系的研究[J]. 武汉植物学研究, 9(3): 239-246. |
彭少麟, 王伯荪. 1983. 广东亚热带森林群落物种多样性[J]. 生态科学, (2): 98-104. |
单人驊, 刘方勋. 1964. 安徽大别山的植被及其地理分布纪要[J]. 植物生态学及地植物学丛刊, 2(1): 93-102. |
吴承祯, 洪伟, 陈辉, 等. 1996. 万木林中亚热带常绿阔叶林种多样性研究[J]. 福建林学院学报, 16(1): 33-37. |
谢晋阳, 陈灵芝. 1994. 暖温带落叶阔叶林的乔木层物种多样性特征[J]. 生态学报, 14(4): 334-337. |
谢中稳, 吴国芳. 1993. 安徽大别山多枝尖山区植物区系的研究[J]. 华东师范大学学报, 自然科学版, (1): 102-110. |
谢中稳. 1994. 安徽鹞落坪自然保护区植物区系基本特征的研究[J]. 安徽农业大学学报, 21(4): 507-512. |
谢中稳, 杜国华, 张尽忠, 等. 1994. 安徽省鹦落坪自然保护区植物区系的数量分析[J]. 生物数学学报, 9(4): 240-246. |
谢中稳, 吴国芳. 1995. 安徽省鹞落坪自然保护区植被类型及其分布[J]. 华东师范大学学报, (3): 93-102. |
徐慧, 钱谊, 彭补拙, 等. 2002. 鹞落坪国家级自然保护区生态评价研究[J]. 农业环境保护, 21(4): 360-364. |
Getzin S, Wiegand T, Wiegand K, et al. 2008. Heterogeneity in? uences spatial patterns and demographics in forest stands[J]. Journal of Ecology, 96: 807-820. DOI:10.1111/jec.2008.96.issue-4 |
Klinka K H Y H, Chen Q, Wang L, et al. 1996. Forest canopies and their influence on the cover of understory vegetation in early-weral stands on west Vancouver Islands[J]. Northwest Science, 70: 193-200. |
Maltez-Mouro S, García LV, Maraón T, et al. 2005. The combined role of topography and overstorey tree composition in promoting edaphic and floristic variation in a Mediterranean forest[J]. Ecological Research, 20(6): 668-677. DOI:10.1007/s11284-005-0081-6 |
Messier C, Parent S, Bergeron Y. 1998. Effects of overstory and understory vegetation on the understory light environment in mixed boreal forests[J]. Journal of Vegetation Science, 9: 511-520. DOI:10.2307/3237266 |
Ricklefs R E. 2004. A comprehensive framework for global patterns in biodiversity[J]. Ecology Letters, 7: 1-15. DOI:10.1046/j.1461-0248.2003.00554.x |