 2010, Vol. 46
2010, Vol. 46文章信息
- 李宏群, 廉振民, 陈存根
- Li Hongqun, Lian Zhenmin, Chen Cungen
- 陕西黄龙山自然保护区褐马鸡冬季栖息地的选择
- Wintering Habitat Selection by Brown-Eared Pheasant (Crossoptilon mantchuricum) in Huanglong Mountains Nature Reserve, Shaanxi Province, China
- 林业科学, 2010, 46(6): 102-106.
- Scientia Silvae Sinicae, 2010, 46(6): 102-106.
-
文章历史
- 收稿日期:2009-03-28
- 修回日期:2009-08-04
-
作者相关文章



2. 长江师范学院生命科学系 重庆 408100;
3. 西北农林科技大学资源与环境学院 杨凌 712100
2. Department of Life Science, Yangtze Normal University Chongqing;
3. College of Resource and Environmental Science, Northwest A & F University Yangling 712100
褐马鸡(Crossoptilon mantchuricum)为我国特有珍稀鸟类,国家一级重点保护野生动物,是世界易危鸟类之一,被我国列为濒危物种(郑光美等,1998;IUCN,2000)。其目前主要分布于山西吕梁山、陕西黄龙山、河北小五台山和北京东灵山等地的局部地区(卢欣等,1998),且由于地理屏障和自然植被的破坏,其分布区已被严重分割成3个区域,分别形成3个地理种群,即山西吕梁山脉的中部种群、河北与北京地区的东部种群和陕西的西部种群(Zhang et al., 2002)。数学模型是研究动物栖息地选择的重要方法之一,这些模型从线性回归模型逐步发展到多元曲线回归模型(Morrison et al., 1987;Guisan et al., 2000),其中Logistic回归模型是最常见的模型之一(Pearce et al., 2000;McGrath et al., 2003)。逻辑斯蒂回归模型是对二态变量进行回归分析的标准方法,由于其结果的直观性和预测的可靠性,被逐渐用于鸟类栖息地选择研究中(李文军等,2000;McGrath et al., 2003;张国钢等,2005;徐基良等,2006;江红星等,2009)。过去对褐马鸡栖息地的研究主要集中微环境或者在山西境内(张国钢等,2003;2005; 李宏群等, 2007)。本文首次通过系统取样运用逻辑斯蒂回归模型方法,建立越冬栖息地模型,对褐马鸡西部种群冬季栖息地的选择进行了分析,希望研究结果为该保护区的管理提供科学依据。
1 研究地区陕西延安黄龙山褐马鸡自然保护区(35°28′—36°02′N,109°38′—110°12′E)位于延安市的黄龙、宜川2县交界处,地处陕北黄土高原东南部的黄龙山腹地,南北宽39.5 km,东西长36.6 km,垂直分布范围在海拔962.6~1 783.5 m,相对高差820.9 m,总面积1 942 km2,林地面积为1 682 km2。研究地区设在保护区的核心区北寺山林区,该处境内人口密度较小,交通闭塞,地形起伏,沟壑纵横。四季分明,年平均气温8.6 ℃,极端最低气温为-22.5 ℃,最高气温为36.7 ℃,年平均降雨611.8 mm,多集中在7—9月,年蒸发量856.5 mm,属于大陆性暖温带半湿润气候类型。有关保护区的植被见文献(李宏群等,2007)。
2 研究方法 2.1 数据收集在2006年11月—2007年1月对陕西黄龙山林区的褐马鸡种群进行了调查,采用样带法对整个研究区域进行系统取样调查。根据以往对白马鸡的研究(贾非等,2004),主要对以下16种生境变量进行调查:海拔、东经、北纬、坡度、坡向、乔木盖度、乔木高度、乔木胸径、灌木盖度、灌木高度、草本盖度、草本高度、距最近水源距离、距最近乡间土路距离、距最近居民点距离和距最近林边距离。具体方法为:在研究区内,不同海拔每隔每200 m设置一条样带,按每200 m选取1个10 m×10 m的大样方,其内设4个5 m×5 m中样方和5个1 m×1 m小样方,其中大样方用于测量与乔木有关的变量,中样方用于测量与灌木有关的变量,小样方用于测量与草本有关的变量。小样方设置方法,将10 m×10 m样方的每条对角线都四等分,在1/4、1/2和3/4处各取1个1 m×1 m的小样方,共取5个;中样方是把大样方等分。以上灌木和草本变量测量数据的平均值作为该样方相关变量的数值。
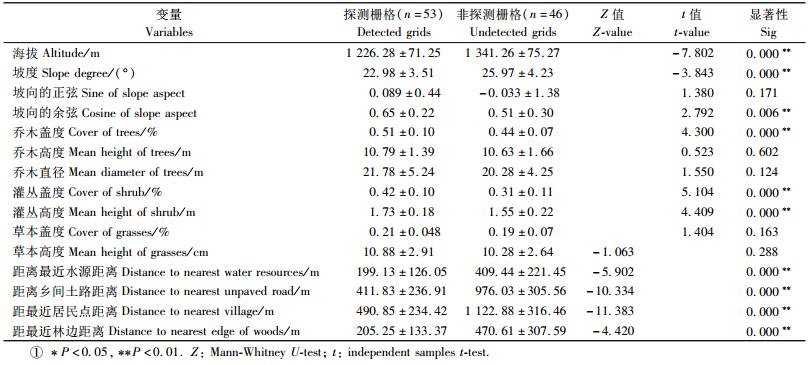
2.2 数学建模 2.2.1 逻辑斯蒂模型逻辑斯蒂模型的一般表达式为π(x)=eg(x)/(1+eg(x)),其中π(x)为物种的出现概率,g(x)=b0+b1x1+b2x2+b3x3+…+bnxn,式中b0为常数,b1,b2,b3…bn为回归系数,x1,x2,x3…xn为变量。在栅格中,有褐马鸡种群出现的赋值为1,定义为探测栅格,反之为非探测栅格,赋值为0。以每个栅格中4个相邻样方环境变量的平均值为该栅格的变量值。对利用组和对照组生境变量的差异进行比较,先用Kolmogorov Smirnov Z检验数据是否符合正态分布。如果原始数据符合正态分布,则使用独立样本的t检验;如果原始数据不符合正态分布,则使用Mann-Whitney U检验。坡向数据属于圆形数据(circular data),取其正弦值和余弦值与其他数据一起分析。差异显著(P < 0.05)的数据进入后续分析。
首先对呈显著差异的变量为自变量,以褐马鸡种群在栅格中的有/无(1/0)为因变量进行单变量逻辑斯蒂回归分析(univariate analysis),P值小于0.30的变量被保留并进入后续分析(McGrath et al., 2003;贾非等,2005;徐基良等,2006)。对保留变量进行相关分析(Spearman双尾相关分析),如果变量间相关系数(|r|>0.70),则对具有生物学意义变量予以保留(贾非等,2005;McGrath et al., 2003)。以保留的变量为自变量,以褐马鸡种群的有无为因变量,采用向前筛选的逐步逻辑斯蒂回归来确定影响褐马鸡栖息地选择的关键因子。
对回归结果计算其AIC及AICc值。AIC或AICc值越小,则该栖息地变量对褐马鸡栖息地选择影响越大;AIC或AICc值均属于Akaike信息标准,现在广泛应用于模型的选择(Pan,2001;Boyce et al., 2002)。当n/K < 40时,一般使用AICC(Burnham et al., 1998;Pan,2001):
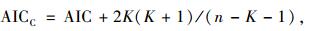
|
式中,K为回归变量的个数+2,n为样本总数。
数据采用Mean±SD表示,其中Mean为算术平均值,SD为标准差。数据处理在SPSS13.0上进行。
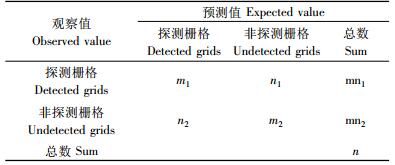
2.2.2 模型检验对所得模型进行Hosmer and Leweshow检验,以此确定模型对因变量变化的判别是否达到显著水平(P>0.05)(Hosmer et al., 1989),同时计算模型的最佳切断点(cut-off point)以及m1,m2,n1,n2,mn1,mn2和n的数值,这些数值用来计算模型对褐马鸡种群对栖息地选择进行预测的准确程度(李文军等,2000)。
|
|
在北寺山447 hm2林区的调查中共发现6个褐马鸡群,平均大小为(7.17±2.86)只。样线共9条,每条样线的平均长度为(2 240±1 031.93)m,根据每个200 m×200 m的栅格是否有褐马鸡的活动痕迹,将栅格分为53个探测栅格和46个非探测栅格。对探测栅格和非探测栅格的变量进行差异性检验(表 2),得到海拔、坡度、坡向的余弦、乔木盖度、灌丛盖度、距离最近水源距离、距离最近乡间土路距离、距最近居民点距离以及距最近林边距离等呈显著差异。经过逻辑斯蒂回归的单变量分析,变量海拔、坡度、坡向的余弦、乔木盖度、灌丛盖度、距离最近水源距离、距离最近乡间土路距离、距最近居民点距离、距最近林边距离等进入后续分析(P < 0.3)。使用Spearman correlation对这些剩余差异显著变量进行相关性分析,发现海拔与距离最近水源距离、距离最近乡间土路距离、距最近居民点距离相关性性显著(|r|>0.70),实际观察发现,冬季褐马鸡群体选择栖息地时,对水源、乡间土路、居民点的选择不明显,因此该变量被筛选掉。以最后剩余变量作为自变量,以褐马鸡探测栅格和非探测栅格作为因变量采用向前筛选的逐步逻辑斯蒂回归分析,得到3个回归模型(表 3)。根据各模型的AICC值(表 2),最终选定模型Ⅲ为最佳模型,其表示为:
|
|
|
|
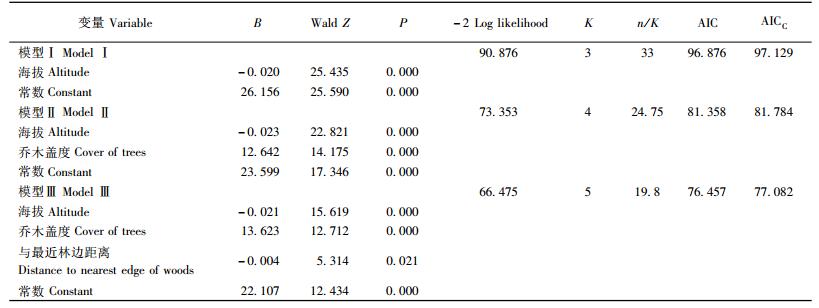
π(x)=eg(x)/(1+eg(x)),g(x)=22.107-0.004×距最近林边距离+13.623×乔木盖度-0.021×海拔。
3.2 模型检验Hosmer and Leweshow检验表明,该模型Ⅲ对因变量变化的判别达到显著水平(χ2=10.344,df=8,P=0.242),根据切断点,m1,m2,n1,n2,mn1,mn2和n等参数的定义及其计算方法,得到模型的最佳切断点为0.63(图 1),并且m1= 48,n1=5,mn1= 53,n2= 10,m2=36,mn2=46,n=99,代表整个模型预测准确度程度的CT[(m1+m2)/n]= 84.85%;代表探测栅格预测正确程度的CP(m1/ mn1)= 90.57%;代表非探测栅格预测准确程度CA(m2/mn2)= 78.26 %。以上结果表明,该模型对褐马鸡冬季栖息地选择具有较高的预测准确性。

|
图 1 冬季褐马鸡群在不同切断点下对栖息地选择模型的正确预测值 Figure 1 Correctly classified ratio of habitat selection model under different cut-off points for activity probability of brown eared-pheasent in winter |
冬季气候寒冷,食物缺乏,对动物的生存很不利,同时植物凋谢,积雪覆盖,隐蔽条件极差,更易遭受天敌的捕食。与此相适应,许多鸟类,尤其是大多数雉类以集群方式越冬(贾陈喜等,1999)。褐马鸡作为地栖生活的大型森林鸟类,与大多数雉类一样,越冬期间常形成大小不同的群体,并倾向于到郁闭度较高的针叶林和针阔混交林中共同觅食(张国钢等,2003; 2005)。对每一个群体来说,由于它们扩散能力较差,一般活动区域比较固定,通过对黄龙山林区褐马鸡活动范围的观察,根据群体大小以及活动范围,可以把观察到的褐马鸡种群可分为北寺山种群、西洼种群、麻子山种群、松树洼狼沟种群、梅家山种群和松树洼后山种群,平均大小为7.17±2.86。这种集群大小与其选择栖息地相适应的。
地形因素和植被结构是决定雉鸡类在某一尺度上栖息地选择的关键因子(徐基良等,2006;康明江等,2008;丛培昊等,2008)。通过逻辑斯蒂回归建立的数学模型为:π(x)=eg(x)/1+eg(x),g(x)=22.107-0.004×距林边距离+13.623×乔木盖度-0.021×海拔。该模型对褐马鸡冬季栖息地选择具有较高的预测准确性,达到84.85%。该模型反映出褐马鸡种群冬季对栖息地选择3个最重要的生态因子为海拔、乔木盖度和距最近林边的距离。种群出现概率与乔木盖度成正相关,与海拔和最近林边距离成负相关。乔木盖度是褐马鸡冬季重要的环境因子,这一点在白马鸡(Crossoptilon crossoptilon)、白冠长尾雉(Syrmaticus reeviesii)和蓝马鸡(Crossoptilon auritum)冬季栖息地选择中同样存在(贾非等,2004;刘振生等,2005;徐基良等,2006)。刘焕金等(1991)和张国钢(2005)的研究也表明,针叶林是褐马鸡冬季经常利用的栖息地类型,褐马鸡常常聚集在一起在高大的树木下取食松子(刘焕金等,1991;张国钢等,2005)。在冬季,由于气温下降,天气寒冷等原因,落叶阔叶林季节性大面积的落叶,造成阔叶林中的郁闭度降低,隐蔽性较差,致使褐马鸡种群移向山地针叶林中,在黄龙山腹地林区主要以油松为主,与其他林型相比,以油松(Pinus tabulaeformis)为主的针叶林在冬季的郁闭度较高,且林下有大量地油松种子,这样既满足褐马鸡隐蔽条件,具有较低的捕食压力,又能满足其对食物的需要,张国钢等报道冬季褐马鸡主要取食松子和草根(张国钢等,2005),这造成褐马鸡在冬季必须选择乔木盖度较大的针叶林。同时,笔者也发现松树林中往往积雪较早融化,说明此林中温度较高,可能也是其选择针叶林的重要原因。因此,栖息地的选择与乔木盖度成正相关。这也进一步证实张国钢等提出的大尺度范围内针叶林面积起关键作用的结论(张国钢等,2005)。此外,林边常有一些阔叶乔木如漆树(Toxicodendron vernicifluum)、野山楂(Crataegus pinnatifida)以及核桃楸(Juglans mandshurica)等,这些乔木的种子或者果实,往往成为褐马鸡冬季喜爱的食物;一些灌丛在森林边缘发育良好如中国沙棘(Hippophae rhamnoides)、连翘(Forsythia suspense)、黄刺玫(Rosa xanthina)、水栒子(Cotoneaster multiflorus)等,其常常具有丰富的浆果以及根部的韧皮部;还有草本植物如独角莲(Typhonium giganteum)的块茎,也是褐马鸡冬季的主要食物,当其觅食时,如果遇到危险,其可以快速逃进针叶林,所以野外常看见林边有大量褐马鸡觅食的痕迹。
褐马鸡在冬季都喜欢在低海拔下坡位觅食(刘焕金等,1991)。在黄龙山,冬季褐马鸡多选择低海拔和下坡位,由于降雪和温度较低,高海拔和山体上部都已接冻,给褐马鸡活动造成不便,山体下部温度较高,冰雪融化比较早,迫使褐马鸡在深冬向低海拔山体下部移动。还有,冬季大雪封山以后,进山的人数明显减少,这为褐马鸡山下活动创造了有利条件,结果褐马鸡常常下移到路边田地活动,因此,褐马鸡冬季栖息地与海拔负相关。
丛培昊, 郑光美. 2008. 四川老君山地区红腹角雉的夜栖行为和夜栖地选择[J]. 生物多样性, 16(4): 332-338. |
贾陈喜, 郑光美, 周小平. 1999. 卧龙自然保护区雪雉的社群组织[J]. 动物学报, 45(2): 135-142. |
贾非, 王楠, 郑光美. 2004. 白马鸡冬季群体栖息地选择的经验模型[J]. 北京师范大学学报:自然科学版, 40(4): 524-530. |
贾非, 王楠, 郑光美. 2005. 冬季白马鸡群体夜栖地特征分析[J]. 生态学杂志, 24(2): 153-158. |
江红星, 刘春悦, 钱法文, 等. 2009. 基于3S技术的扎龙湿地丹顶鹤巢址选择模型[J]. 林业科学, 45(7): 76-83. |
康明江, 郑光美. 2007. 白腹锦鸡(Chrysolophus amherstiae)的夜栖地选择[J]. 生态学报, 27(7): 2929-2934. |
李宏群, 廉振民, 陈存根, 等. 2007. 陕西黄龙山林区褐马鸡春季觅食地选择的研究[J]. 动物学杂志, 42(3): 61-67. |
李文军, 王子健. 2000. 丹顶鹤越冬栖息地数学模型的建立[J]. 应用生态学报, 11(6): 839-842. |
刘振生, 曹丽荣, 李志刚, 等. 2005. 贺兰山蓝马鸡越冬期栖息地的选择[J]. 动物学杂志, 40(2): 38-43. |
刘焕金, 苏化龙, 任建强. 1991. 中国雉类——褐马鸡[M]. 北京: 中国林业出版社.
|
卢欣, 郑光美, 顾滨源. 1998. 马鸡的分类、分布及演化关系的初步探讨[J]. 动物学报, 44(2): 131-137. |
徐基良, 张晓辉, 张正旺, 等. 2006. 白冠长尾雉越冬期栖息地选择的多尺度分析[J]. 生态学报, 26(7): 2061-2067. |
张国钢, 张正旺, 郑光美, 等. 2003. 山西五鹿山褐马鸡不同季节的空间分布与栖息地选择研究[J]. 生物多样性, 11(4): 303-308. |
张国钢, 郑光美, 张正旺, 等. 2005. 山西芦芽山褐马鸡越冬栖息地选择的多尺度研究[J]. 生态学报, 25(5): 952-957. |
郑光美, 王岐山. 1998. 褐马鸡//王松. 中国濒危动物红皮书(鸟类). 北京: 科学出版社, 242-243.
|
Boyce M S, Vernier P R, Nielsen S E, et al. 2002. Evaluating resource selection functions[J]. Ecological Modelling, 157: 281-300. DOI:10.1016/S0304-3800(02)00200-4 |
Burnham K P, Anderson D R. 1998. Model selection and inference: a practical information-theoretic approach[M]. New York: Springer-Verlog: 76-81.
|
Guisan A, Zimmermann N E. 2000. Predictive habitat distribution models in ecology[J]. Ecological Modelling, 135: 147-186. DOI:10.1016/S0304-3800(00)00354-9 |
Hosmer D W, Lemeshow S. 1989. Applied Logistic Regression[M]. New York: John Wiley and Sons: 86-89.
|
IUCN. 2004. IUCN Red List of Threatened species. http://www.redlist.org.[2004-06-16].
|
McGrath M T, DeStefand S, Riggs R A. 2003. Spatially explicit inflences on northern goshawk nesting habitat in the interior Pacific Northwest[J]. Wildlife Monographs, 154: 1-63. |
Morrison M L, Timossi I C, With K A. 1987. Development and testing of linear regression models predicting bird-habitat relationships[J]. Journal of Wildlife Management, 51: 247-253. DOI:10.2307/3801663 |
Pan W. 2001. Akaike' s information criterion in generalized estimating equations[J]. Biometrics, 57(1): 120-125. DOI:10.1111/j.0006-341X.2001.00120.x |
Pearce J, Ferrier S. 2000. Evaluating the predictive performance of habitat models developed using logistic regression[J]. Ecological Modelling, 133: 225-245. DOI:10.1016/S0304-3800(00)00322-7 |
Zhang Z W, Zheng G M, Zhang G G, et al. 2002. Distribution and Population Status of Brown eared-Pheasant in China[M]. UK: World Pheasant Association: 91-96.
|