 2010, Vol. 46
2010, Vol. 46文章信息
- 王雷, 尹增芳, 尤禄祥, 李建银, 于明祥, 徐棉芬
- Wang Lei, Yin Zengfang, You Luxiang, Li Janyin, Yu Mingxiang, Xu Mianfe
- 外源Ca2+对南林895杨扦插苗生长及稳定同位素δ13C, δ15N动态分布的影响
- Effects of Exogenous Ca2+ on Growth and Dynamic Distribution of Stable Isotope δ13C, δ15N of Populus×euramericana 'Nanlin 895' Cuttings
- 林业科学, 2010, 46(6): 51-57.
- Scientia Silvae Sinicae, 2010, 46(6): 51-57.
-
文章历史
- 收稿日期:2009-03-23
- 修回日期:2009-11-07
-
作者相关文章



钙(Calcium)是植物必要的营养元素之一,它在植物发育过程的各种生理活动中扮演重要的角色。Ca2+能够维持植物生物膜的稳定性,对植物细胞分裂和分化、器官形态建成、各种生化反应过程都起到关键的调节作用(潘瑞炽等,2000)。在植物个体发育过程中,外源Ca2+可明显地促进植物的营养生长、提高植株产量,这一结论在莴苣(Lactuca sativa)(范双喜等,2002)、青檀(Pteroceltis tatarinowii)(洑香香等,2004)、大豆(Glycine max)(王晶英等,2004)、番茄(Solanum lycopersicum)(董彩霞等,2005)、马铃薯(Solanum tuberosum)(辛建华等,2008)、东南景天(Sedum alfredii)(黄化刚等,2008)等植物中得到进一步的证实。
现有资料表明,植物δ13C可以很好地反映植物的光合、呼吸、蒸腾、水分利用率等多种指标(Condon et al., 2002; Rebetzke et al., 2002; 丁明明等,2006;赵凤君等,2005),δ15N可以作为大气沉氮、植物营养状况、氮利用等的指示剂(Michopoulos et al., 2004; 李德军等, 2003; Hogberg, 2007; Guerrieri et al., 2009),从而揭示植物正常的生长发育状况。尹伟伦等(2007)研究发现δ13C不仅可作为评价不同杨树无性系整株水平长期水分利用效率的良好指标,而且还可能被用于预测和评价良好水分条件下杨树的生长潜力。已经证实δ13C与产量的关系受到环境条件、作物的物候期和用来分析δ13C的植物器官的影响(Araus et al., 2003; Royo et al., 2008), 但在同样的生长条件下杨树器官间稳定同位素δ13C及其δ15N变化规律的研究尚未见报道。本试验以南林895杨(Populus×euramericana 'Nanlin 895')为试验材料,进行外源Ca2+叶面累积喷施处理, 采用形态检测与稳定同位素分析技术相结合的方法,探讨外源Ca2+作用下南林895杨扦插苗生物量规律性的变化,以及生长过程中稳定同位素δ13C,δ15N在不同营养器官间分馏的分布趋势,为杨树生产实践提供理论技术指导。
1 材料与方法 1.1 试验材料3月5日选取土肥一致的地块,将南林895杨插穗按照25 cm×40 cm的株行距进行扦插,试验总体样本数共计500株。3个月后选择苗高(50 cm左右)、健壮度一致及无病虫害的苗木作为试验材料。
1.2 外源Ca2+叶面累积喷施处理以CaCl2溶液喷施扦插苗,外源Ca2+喷施质量浓度设为0,2,20,100,200,300 mg·L-1等6个水平。每种处理重复喷施30株,每隔15天喷施1次,共计喷施3次。喷施时间选在无风晴朗的时间段进行,喷施强度至叶的正反面滴水为止。
1.3 指标测定 1.3.1 株高、地径株高、地径分别在每次喷施15天后进行测定,分别从每种处理中选取30株能够代表各水平总体生长趋势的扦插苗做为试验样本,用皮尺测定植株的高度,用游标卡尺测定植株的地径。
1.3.2 生物量累积喷施3次外源Ca2+后,7月20日分别从各处理中随机选择10株扦插苗,将每一植株的根、茎、叶分别采集。并将采集的样品放入70 ℃烘箱中烘干(先110 ℃杀青15 min),用分析天平分别称量出根、茎、叶等器官的生物量(干质量),并计算出单株植物的生物总量。
1.3.3 δ13C及δ15N值每个处理采集10株样本,按根、茎、叶分别于70 ℃烘箱中烘干,并用高速万能磨碎机将其磨碎至粉末状,过40目筛。按照不同营养器官充分混合后,随机抽取一定量的样品(1.5 g)进行δ13C及δ15N值测定(主机型号:DELTAplus XP;精度:δ13C<0.1‰ δ15N<0.2‰;每隔12个样品放入2个实验室标准物质(glycine和urea)。
1.4 数据处理应用Excel 2003、Spass 11.5软件进行数据分析处理。
2 结果与分析 2.1 外源Ca2+处理下杨树扦插苗形态与生物量的变化 2.1.1 不同质量浓度Ca2+处理对扦插苗株高的影响从图 1可以看出,不同质量浓度的外源Ca2+处理南林895杨扦插苗,其株高在第15,30,45天的变化趋势基本一致;即在2,20,100,200 mg·L-1质量浓度处理下呈递增趋势,质量浓度为300 mg·L-1时,株高增长呈逐渐下降的趋势,而且这种变化趋势以连续喷施3次后的效果最为明显。Duncan多重比较结果表明:首次进行外源Ca2+处理后,2 mg·L-1质量浓度的Ca2+处理的扦插苗,其株高与对照(CK)无显著差异;而100,200,300 mg·L-1处理扦插苗高生长均优于对照(CK),且达到显著水平,其中200 mg·L-1处理,达到极显著水平。累积喷施Ca2+第30天后,扦插苗的生长明显加快,随着Ca2+质量浓度的升高,扦插苗高生长均优于对照(CK)。但2, 20 mg·L-1质量浓度Ca2+处理与对照(CK)相比,其株高增长量没有显著差异,而100,200 mg·L-1 Ca2+质量浓度处理的株高增长与对照(CK)相比差异达到显著水平1,分别比对照增长4.87%和5.79%。与前2次处理一样,累积喷施第45天后,扦插苗高生长均优于对照(CK)。其中以200 mg·L-1 Ca2+质量浓度处理最佳,净增长达到137.90 cm(CK为120.10 cm),与对照(CK)相比差异达到极显著水平。100, 300 mg·L-1 Ca2+质量浓度处理的扦插苗株高与对照(CK)均达到显著水平,但2,20 mg·L-1 Ca2+质量浓度处理与CK均无显著差异(表 1)。说明低质量浓度外源Ca2+水平对南林895杨扦插苗株高生长的累积效应不明显,而较高质量浓度外源Ca2+处理对扦插苗的高生长具有促进作用,其中以200 mg·L-1 Ca2+质量浓度处理最佳。但过高质量浓度的外源Ca2+延缓了植株的高生长。

|
图 1 累积喷施外源Ca2+后南林895杨扦插苗株高的动态变化 Figure 1 Dynamic changes of Populus×euramericana 'Nanlin 895' cuttings plant height in multi-spraying exogenous Ca2+ |
|
|
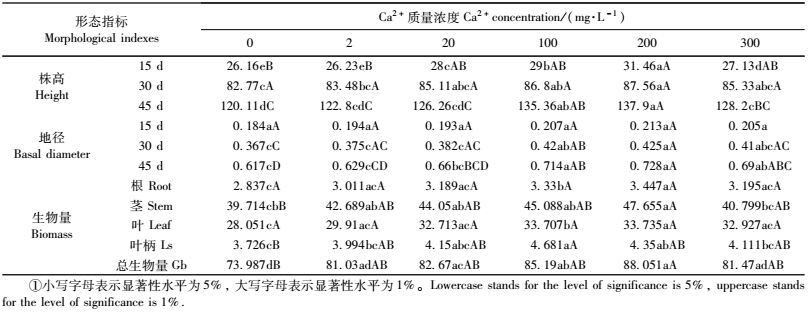
地径的增长趋势与株高生长情况类似(图 2;表 1)。不同质量浓度的外源Ca2+均能促进杨树扦插苗地径的生长。首次喷施Ca2+后,南林895杨地径的生长变化不明显,与对照(CK)无显著差异性。累积喷施Ca2+第30天后,扦插苗地径生长加快,200 mg·L-1质量浓度Ca2+处理的扦插苗的地径生长最好,达到0.425 cm(CK为0.367 cm),与对照(CK)相比差异达到极显著水平,其他Ca2+质量浓度处理下扦插苗地径增长也达到显著水平。说明多次喷施后,外源Ca2+的累积效应开始发生作用。第3次喷施外源Ca2+后,扦插苗地径的净增长量在各Ca2+质量浓度处理间均有不同程度的差异性。100,200,300 mg·L-1质量浓度Ca2+处理与对照(CK)达到极显著水平;以Ca2+质量浓度200 mg·L-1处理地径净增长量最大,达到0.728 cm(CK为0.617 cm);2,20 mg·L-1 Ca2+质量浓度处理可以促进扦插苗地径的生长,但与对照(CK)相比仍无显著性差异。

|
图 2 累积喷施外源Ca2+后南林895杨扦插苗地径的动态变化 Figure 2 Dynamic changes of Populus×euramericana 'Nanlin 895'cuttings Basal diameter in multi-spraying exogenous Ca2+. |
3次累积喷施Ca2+后,统计分析了南林895杨扦插苗“源”“库”器官间生物量的变化情况,结果表明:随着外源Ca2+质量浓度的升高,植株各营养器官间的生物量均高于对照(CK),总生物量以200 mg·L-1 Ca2+质量浓度处理最佳,达到88.05 g(CK为73.99 g),与对照(CK)相比差异极显著,而与其他处理的差异性也达到显著水平。但300 mg·L-1 Ca2+质量浓度处理的植株生物量虽然优于对照,但与200 mg·L-1 Ca2+质量浓度处理相比呈明显下降趋势;叶、根、茎的生物量以200 mg·L-1质量浓度的Ca2+最佳,与CK相比差异达到极显著水平,净增长量分别为33.74 g(CK为28.05 g),3.45 g(CK为2.90 g)、47.66 g(CK为39.71 g)。说明较低质量浓度的外源Ca2+(2, 20 mg·L-1)对南林895杨扦插苗生物量的增长促进作用不明显,而过高质量浓度的外源Ca2+(300 mg·L-1)虽然对生物量的增长具有一定的促进作用,但效果明显低于最佳质量浓度(200 mg·L-1)的外源Ca2+处理(图 3,表 1)。

|
图 3 累积喷施外源Ca2+后南林895杨扦插苗“源”“库”器官生物量的变化 Figure 3 Biomass changes of "sources" and "sink" organ of Populus×euramericana 'Nanlin 895' cuttings in multi-spraying exogenous Ca2+ R根;Ls叶柄;L叶;S茎;G总生物量。 R:root; Ls:leaf stalk; L:leaf; S:stem; G: gross biomass. |
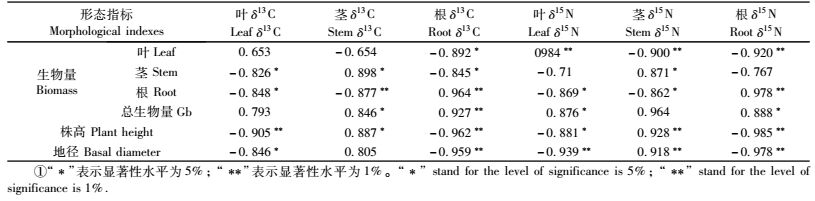
不同质量浓度的外源Ca2+均能不同程度地影响南林895杨扦插苗营养器官碳(C)、氮(N)的分馏状况。随着Ca2+质量浓度的升高,叶、茎、根中δ13C, δ15N值在南林895杨扦插苗营养器官中均呈现下列规律性的变化:δ13C值的变化规律为;叶﹤茎﹤根(图 4)。即植株“源”器官δ13C值明显低于“库”器官,说明稳定同位素C较容易在非光合器官(茎和根)中富集。δ15N值的变化规律为:叶﹤根﹤茎(图 5)。植株“源”器官比“库”器官具有较高的δ15N值,说明在外源Ca2+处理下,N素的分馏发生了相应的变化,“源”器官较易发生同位素N的分馏,而“库”器官更容易富集同位素N。与δ13C不同的是,根比茎更容易发生同位素N的分馏。与对照(CK)相比较,不同质量浓度外源Ca2+处理均能不同程度地提高南林895杨扦插苗的δ13C,δ15N值,且最大值均出现在200 mg·L-1 Ca2+质量浓度的处理中。δ13C和δ15N值在茎中分别比对照高29.5‰和546.9‰;在叶中分别比对照高107.1‰和467.8‰;在根中分别比对照高107.4‰和593.9‰。但是,当外源Ca2+质量浓度达到300 mg·L-1时,δ13C, δ15N的值呈现下降的趋势。

|
图 4 不同质量浓度Ca2+处理下南林895杨扦插苗营养器官δ13C值的动态变化 Figure 4 δ13C value dynamic changes of Populus×euramericana 'Nanlin 895' cuttings in different Ca2+ concentration |

|
图 5 不同质量浓度Ca2+处理下南林895杨扦插苗营养器官δ15N值的动态变化 Figure 5 δ15N value dynamic changes of Populus×euramericana 'Nanlin 895' cuttings in different Ca2+ concentration |
相关性分析结果表明,外源喷施Ca2+可以影响南林895杨扦插苗各营养器官光合同化物的分配。从表 2可以看出,经不同质量浓度外源Ca2+处理的南林895杨扦插苗,株高、地径与不同营养器官δ13C,δ15N值有一定的相关性。扦插苗“源”器官δ13C,δ15N值与“库”器官生物量呈负相关,而与“源”器官生物量成正相关;“库”器官δ13C,δ15N值与“源”器官生物量呈负相关,而与“库”生物量呈正相关性。“源”器官(叶)δ13C值与株高达到极显著负相关,与地径达到显著负相关,“库”器官茎δ13C值与株高达到显著正相关,而与地径相关不明显,“库”器官根与株高和地径均为负相关;“源”器官(叶)δ15N值与株高、地径呈负相关;“库”器官茎δ15N值与地径、株高均达到极显著相关,而“库”器官根δ15N与株高和地径则呈极显著负相关。这说明不同营养器官的δ13C,δ15N的动态变化可以反映南林895杨扦插苗“源”、“库”间光合同化物积累与再分配格局对外源Ca2+质量浓度的响应。
|
|
适当质量浓度的Ca2+对植物生长具有促进作用。试验证明,外源Ca2+能够调节番茄幼苗的生长,在喷施适宜质量浓度Ca2+条件下,植株幼苗的根系活力、叶绿素含量等生理指标均显著增加(董彩霞等,2005)。在水培试验中,加入适量质量浓度的Ca2+,1年生青檀苗高生长量明显高于对照(洑香香等,2004);王晶英等(2004)在大豆研究中也得出类似结论,一定质量浓度外源Ca2+能够平衡大豆植株对N素的均衡利用,从而促进植株的营养生长。Ca2+的这种促进作用一方面是由于它是植物必要的营养元素,另一方面是由于Ca2+能够作为植物的第二信使在植物各种生理生化活动中起调节作用,从而促进植物的营养生长。不同质量浓度外源Ca2+处理均能使南林895杨扦插苗的株高和地径有不同程度的增长。低质量浓度外源Ca2+的促进作用不明显,而较高质量浓度的外源Ca2+处理对扦插苗的株高、地径生长具有明显的促进作用。但随着Ca2+质量浓度的增高,这种促进效应逐渐减缓。植物根系的生长与K+吸收有关(Cramer et al., 1986),过高质量浓度Ca2+会抑制根系的生长,从而减少了根系对K+的吸收。另外由于Ca2+与Mg2+间存在离子拮抗现象, Ca2+质量浓度过高会影响Mg2+的吸收,从而影响植物对养分的均衡吸收,这一结果在莴苣(Lactuca sativa)试验中得到证实(范双喜等,2002)。
Ca2+作为植物必要的营养元素,不仅可以促进植株的营养生长,还可以提高植株的生物产量。在外源喷施Ca2+作用下,不同质量浓度外源Ca2+处理均能促进杨树扦插苗生物量的提高。低质量浓度外源Ca2+的促进作用不明显, 而适量质量浓度的Ca2+对植株生物产量的提高具有促进作用。辛建华等(2008)研究发现,一定外源钙水平可以影响马铃薯块茎的质量,适当提高外源钙的水平可以明显促进马铃薯块茎的膨大。黄化刚等(2008)的试验也得出类似结论,随着营养液中Ca2+质量浓度的增加, 东南景天(Sedum alfredii)总的生物量有所增加, 且根系和地上部干物质量均与Ca2+质量浓度呈正相关。研究者在小麦(Triticum aestivum)的试验中还发现,增加营养液中的Ca2+质量浓度可以明显改善小麦幼苗的生长状况, 增强对营养物质的利用, 促进生物量的积累(王志强等,2008)。此外,Ca2+可以提高青檀、大豆、花生(Arachis hypogaea)(洑香香等,2004;王晶英等,2004;吴文新等,2001)植物生物量的研究也屡有报道。但是,Ca2+质量浓度并非越高越好,而是在一个合适的质量浓度范围内才能有效地发挥作用。在本项研究中,较高质量浓度的Ca2+(300 mg·L-1)虽然对生物量的增长仍具有一定的促进作用,但作用的效果明显低于最佳质量浓度的Ca2+处理。这说明,过高质量浓度的Ca2+降低了对植物生长的促进效应。
3.2 稳定同位素δ13C, δ15N的动态分布对外源Ca2+作用的响应在外源Ca2+作用下,南林895杨营养器官的δ13C值在-28.856‰~-30.169‰之间变化,基本规律为:叶﹤茎﹤根,与对照相比δ13C值均有所提高。这与洪伟等(2008)对桉树(Eucalyptus spp.)的研究结果一致。叶片中的δ13C值要低于植物其他光合器官及其非光合器官(Nilsen et al., 1997),即植物“源”器官(叶)δ13C值明显低于“库”器官(茎和根)。Badeck等(2005)研究发现,植物叶片平均δ13C值要比茎干及根系分别低0.96‰和1.91‰。同样地,南林895杨扦插苗“库”器官比“源”器官更容易富集稳定同位素C。并且在不同质量浓度外源Ca2+处理下,南林895杨扦插苗不同营养器官δ13C值均比对照(CK)要高,因此推测喷施Ca2+可以增加叶片细胞内的Ca2+质量浓度,促使细胞液质量浓度增加,水势降低,故气孔开放程度加大,从而促进了南林895杨叶片光合作用。研究资料表明,光合作用是导致稳定碳同位素分馏的主要因素,这种分馏效应导致了不同组织器官间的δ13C值差异(陈世苹等, 2002; Badeck et al., 2005)。因此,外源Ca2+间接地成为了南林895杨扦插苗各营养器官δ13C值动态变化的影响因子之一。李明财等(2007)对高山林线急尖长苞冷杉(Abies georgei)的稳定碳同位素分析得出,植株各器官δ13C值分布情况为,叶﹤根﹤枝干;与本试验茎δ13C值小于根的结果不一致,这可能是由于不同物种δ13C分布不同及其不同树龄δ13C分布产生差异的结果(冯虎元等,2000)。南林895杨扦插苗在早期生长过程中,根系的形态建成趋势强于茎干,导致碳素分馏程度根大于茎的现象。而导致南林895杨叶片δ13C值偏低的原因,可能由于“源”器官(叶)将所合成的碳水化合物转运到茎和根等部位,使得叶片中的碳水化合物积累减少,因此叶片中稳定同位素δ13C值较低,这一结论与Francey等(1985)的假设一致。
与δ13 C值一样,南林895杨营养器官δ15N值的时空分布也有显著的差异性。外源Ca2+作用下杨树各器官的δ15N值主要在-0.575‰~-2.153‰之间变化,植株各器官δ15N值的变化规律为:叶﹤根﹤茎。说明植株“库”器官比“源”器官具有较高的δ15N值。Hobbie等(2000)研究提出,氮在植物组织间的运移会使叶片氮同位素发生变化,研究证实氮从根部吸收到植物主体其δ15N值相差可达8‰。与δ13C不同的是,根部δ15N值比茎要小,推测其可能的原因是由于扦插苗在不同生长时期,生长中心发生转移的结果,根部吸收氮素将其转移到植株其他部位。
在不同质量浓度外源Ca2+作用下,南林895杨各营养器官δ15N值均比对照要高。在自然环境条件下,植物体主要从大气和土壤中获取氮源(Foyer et al., 2002)。因此,在植物代谢生理过程中氮素的吸收、同化、利用过程是最关键的内容,而Ca2+对植物幼苗氮素代谢的影响作用主要体现在中间代谢物含量上的变化,即铵离子的含量随着Ca2+质量浓度的加大而增加(王志强等,2008)。Ca2+可以促进硝态氮的吸收, 有助于减少植物中的硝酸盐, 中和植物中的有机酸, 对代谢过程中产生的有机酸有解毒作用(曹恭等,2003)。说明Ca2+在植物吸收氮素的过程中起着重要的调节作用。本项研究结果也显示,在适宜质量浓度外源Ca2+处理下南林895杨不仅营养器官中氮素的富集程度提高,而且其生长性状也得到了进一步的改善,说明Ca2+有利于植物对氮素的吸收,并有效地促进了植物的生长。
曹恭, 梁鸣早. 2003. 钙——平衡栽培体系中植物必需的中量元素[J]. 土壤肥料, (2): 48-49. |
陈世苹, 白永飞, 韩兴国. 2002. 稳定性碳同位素技术在生态学研究中的应用[J]. 植物生态学报, 26(5): 549-560. |
董彩霞, 周健民, 赵世杰, 等. 2005. 外源钙对不同钙敏感型番茄幼苗生理特性的影响[J]. 应用生态学报, 16(2): 267-272. |
丁明明, 苏晓华, 黄秦军. 2006. 欧洲黑杨基因资源稳定碳同位素组成特征[J]. 林业科学研究, 19(3): 272-276. |
范双喜, 伊东正. 2002. 钙素对叶用莴苣营养吸收和生长发育的影响[J]. 园艺学报, 29(2): 149-152. |
冯虎元, 安黎哲, 王勋陵. 2000. 环境条件对植物温度碳同位素组成的影响[J]. 植物学通报, 17(4): 312-318. |
洑香香, 方升佐, 田野, 等. 2004. 钙离子浓度对青檀生长和檀皮质量的影响[J]. 应用生态学报, 15(9): 1502-1506. |
黄化刚, 李廷轩, 张锡洲, 等. 2008. 外源钙离子对东南景天生长及锌积累的影响[J]. 应用生态学报, 19(4): 831-837. |
洪伟, 黄锦湖, 李键, 等. 2008. 不同桉树品种稳定碳同位素研究[J]. 福建林学院学报, 28(3): 193-197. |
李德军, 莫江明, 方运霆, 等. 2003. 氮沉降对森林植物的影响[J]. 生态学报, 23(9): 1891-1900. |
李明财, 罗天祥, 刘新圣, 等. 2007. 高山林线急尖长苞冷杉不同器官的稳定碳同位素组成分布特征[J]. 应用生态学报, 18(12): 2654-2660. |
潘瑞炽. 2000. 植物生理学[M]. 4版. 北京: 高等教育出版社: 30.
|
王晶英, 张兴梅, 李国兰. 2004. 钙对大豆生长及产量的影响[J]. 中国油料作物学报, 26(1): 60-62. |
吴文新, 陈家驹, 周恩生, 等. 2001. 钙硼对花生生长产量和品质的影响[J]. 亚热带植物科学, 30(2): 20-23. |
王志强, 王春丽, 林同保. 2008. 外源钙离子对小麦幼苗氮素代谢的影响[J]. 生态学报, 28(8): 3662-3667. |
辛建华, 李天来, 陈红波. 2008. 外源钙处理对马铃薯块茎重量和数量的影响[J]. 西北农业学报, 17(5): 248-251. |
尹伟伦, 万雪琴, 夏新莉. 2007. 杨树稳定碳同位素分辨率与水分利用效率和生长的关系[J]. 林业科学, 43(8): 15-22. |
赵凤君, 高荣孚, 沈应柏, 等. 2005. 水分胁迫下美洲黑杨不同无性系间叶片δ13C和水分利用效率的研究[J]. 林业科学, 41(1): 36-41. DOI:10.11707/j.1001-7488.20050108 |
Araus J L, Villegas D, Aparicio N, et al. 2003. Environmental factors determining carbon isotope discrimination and yield in durum wheat under Mediterranean conditions[J]. Crop Science, 43(1): 170-180. DOI:10.2135/cropsci2003.1700 |
Badeck F W, Tcherkez G, Nogues S, et al. 2005. Post-photosynthetic fractionation of stable carbon isotopes between plant organs: A widespread phenomenon[J]. Rapid Communications in Mass Spectrometry, 19: 1381-1391. DOI:10.1002/(ISSN)1097-0231 |
Cramer G R, Lauchli A, Epstein E. 1986. Effects of NaCl and CoC/Zon ion activities in complex nutrient solutions and root growth of cotton[J]. Plant Phsiol, 81: 792-797. DOI:10.1104/pp.81.3.792 |
Condon A G, Richards R A, Rebetzke G J, et al. 2002. Improving intrinsic water-use efficiency and crop yield[J]. Crop Sci, 42: 122-131. DOI:10.2135/cropsci2002.1220 |
Francey R J, Giord R M, Sharkey T D, et al. 1985. Physiological influences on carbon isotope discrimination in huon pine (Lagarostrobus franklinii)[J]. Oecologia, 66: 469-481. |
Foyer C H, Noctor G.2002.Photosynthetic nitrogen assimilation: inter-pathway control and signaling//Foyer C H, Noctor G. photosynthetic-nitrogenassimilation and associated carbon and respiratory metabolism. Dordrecht: Kluwer Academic Publishers, 1-22.
|
Guerrieri M R, Siegwolf R T W, Saurer M, et al. 2009. Impact of different nitrogen emission sources on tree physiology as assessed by a triple stable isotope approach[J]. Atmospheric Environment, 43: 410-418. DOI:10.1016/j.atmosenv.2008.08.042 |
Hogberg P. 2007. Nitrogen impacts on forest carbon[J]. Nature, 447: 781-782. DOI:10.1038/447781a |
Hobbie E A, Macko S A, Williams M. 2000. Correlations between foliar δ15N and nitrogen concentrations may indicate plant mycorrhizal interactions[J]. Oecologia, 122: 273-283. DOI:10.1007/PL00008856 |
Michopoulos P, Baloutsos G, Economou A, et al. 2004. Effects of nitrogen deposition on nitrogen cycling in an Aleppo-pine stand in Athens, Greece[J]. Science of the Total Environment, 323: 211-218. DOI:10.1016/j.scitotenv.2003.10.002 |
Nilsen E T, Sharifi M R. 1997. Carbon isotopic composition of legumes with photosynthetic stems from Mediterranean and desert habitats[J]. American Journal of Botany, 84(12): 1707-1713. DOI:10.2307/2446469 |
Royo C, Martos V, Ramdani A, et al. 2008. Changes in yield and carbon isotope discrimination of Italian and Spanish durum wheat during the 20th Century[J]. American Society of Agronomy, 100: 352-360. DOI:10.2134/agronj2007.0060 |
Rebetzke G J, Condon A G, Richards R A, et al. 2002. Selection for reduced carbon isotope discrimination increases aerial biomass and grain yield of rainfed bread wheat[J]. Crop Sci, 42: 739-745. DOI:10.2135/cropsci2002.0739 |