 2010, Vol. 46
2010, Vol. 46文章信息
- 张骏, 葛滢, 江波, 常杰, 袁位高, 朱锦茹, 戚连忠
- Zhang Jun, Ge Ying, Jiang Bo, Chang Jie, Yuan Weigao, Zhu Jinru, Qi Lianzhong
- 浙江省杉木生态公益林碳储量效益分析
- Carbon Storage Efficiency of Cunninghamia lanceolata Ecological Service Forest in Zhejiang
- 林业科学, 2010, 46(6): 22-26.
- Scientia Silvae Sinicae, 2010, 46(6): 22-26.
-
文章历史
- 收稿日期:2009-02-28
- 修回日期:2009-05-19
-
作者相关文章



2. 浙江大学生命科学学院 杭州 310058
2. College of Life Science, Zhejiang University Hangzhou 310058
在当今全球生态环境日益恶化、生态系统日趋失衡的大背景下,作为陆地的主体——森林的生态效益愈发重要,生态公益林的建设成为改善生态环境的重要组成部分。生态公益林与商品林定义相对应,是指人们根据需要而指定的现存或即将营造和改造的不以生产直接的有形产品而以生产生态效益产品为目的的森林(周国逸等,2000)。生态公益林的效益包括生态效益、经济效益和社会效益。生态效益体现在生物量、生产力、水文效应、水土保持、固土保肥、改良土壤、净化大气和保护生物多样性等方面(方奇,2000; 田大伦等,2003),生物量和碳储量是各种生态效益的基础,所以国内外森林生物量和碳储量的研究较多(冯宗炜等,1982; Brown et al., 1999; Cairns et al., 2003; 方精云等,2006; 胡会峰等,2006; Zhang et al., 2007)。
近年来浙江省森林恢复迅速,覆盖率已达59.4%(刘安兴等,2002),其中针叶林占森林面积的85%。为了加强生态环境建设,于2001年全面启动生态公益林建设工程,划归生态公益林面积占全省森林面积的22.8 %。杉木(Cunninghamia lanceolata)是与中国南方环境相适应的重要的速生树种,是南方首要的商品材树种。20世纪50年代杉木造林面积迅速增加(浙江森林编辑委员会,1993)。随着生态公益林建设工程的推进,很多分布在水源涵养林区、生态保护植被恢复区和森林生态治理区的杉木林也被划分为生态公益林。现有已归属为生态公益林的杉木林面积达10.53万hm2,占全省生态公益林面积的12.8 %(刘安兴等,2002)。
本研究探讨浙江省不同年龄杉木优势林和含杉木混交林在生物量和碳储量方面的生态效益,为大面积林型更新提供依据,并为正确评价浙江森林在全球碳平衡中的作用提供部分基础数据。
1 研究区概况浙江省(118°01′—123°10′E, 27°06′—31°11′N)位于中国长江中下游东南沿海地区,长江三角洲南翼,东西宽与南北长相近(约450 km)。省内高山基本集中于西南部,平均海拔800 m。浙江省属于典型的亚热带气候,水热基本同期(浙江森林编辑委员会,1993)。年均气温15.3~18.5 ℃,最低月均气温2.7~7.9 ℃,最高月均气温27.0~29.5 ℃,≥10 ℃年积温4 800~5 800 ℃,全年无霜期225~280天; 年降水量1 000~2 000 mm,以春雨、梅雨、台风雨为主,7—8月有伏旱。土壤类型多以红壤、红黄壤和黄壤为主,还有少量的石灰土、紫色土等,适合于森林群落的生长,故浙江省境内的森林群落类型丰富,植物种类繁多。地带性植被为常绿阔叶林、常绿阔叶和落叶混交林; 除此之外还有落叶阔叶林、杉木林、马尾松(Pinus massoniana)林和竹林等多种植被类型(刘安兴等,2002)。浙江省位于杉木栽培区域中带,栽培历史悠久。
2 研究方法 2.1 样地设置和取样在浙江省23个公益林试点县里选取杉木生态公益林样地67个,其中有年龄的样地46个。样地分布在浙西北、浙中、浙南和浙东南沿海,海拔50~650 m,坡度10~40°,胸径4.3~14.0 cm,年龄为6~30年,群落类型除杉木优势林外还包括杉木-苦槠(Castanopsis sclerophylla)混交林、杉木-木荷(Schima superba)混交林、杉木-苦楝(Melia azedarach)混交林和杉木-拟赤杨(Alniphyllum fortunei)混交林等(黄承才等,2006)。
对选定样地进行群落植被结构、数量特征以及土壤厚度、海拔和坡度等指标的调查。群落层次的划分同方精云等(2004)在中国山地植物物种多样性中采用的方法。各样地面积均为20 m×30 m。对乔木层进行每木检尺,起测胸径为3.0 cm,调查因子为胸径、树高、枝下高、冠幅,被测树木用塑料牌或金属牌编号挂牌; 沿样地对角线设1.0 m宽的下木层调查带,调查每一树种的株数、树高并统计各树种的数量与平均高度; 在样地对角线端部设2个2 m×2 m的草本层小样方,调查小样方内各植物种数量和盖度。本研究的野外调查工作在1999—2000年进行。
2.2 生物量估算在67个杉木样地外围寻找杉木标准木,共伐得52株杉木标准木。根据分层切割法将标准木从底部每2 m截为一段,称鲜质量; 再取其中树干一段和部分侧枝、叶、花和果实,分别称鲜质量,烘干后得干质量,计算各部分含水率再根据各部分总鲜质量推算得各部分总干质量。各部分总干质量相加即得各个杉木标准木的单株生物量(林生明等,1991)。
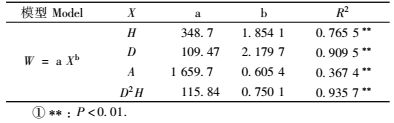
采用维量分析法,对杉木地上生物量(W)分别用株高(H)、胸径(D)和年龄(A)建立回归方程(表 1)。计算得出模型W = 115.84(D2H)0.750 1,相关系数最大(r=0.967 3,P < 0.01),可用来推算样地中未砍伐杉木的生物量。
|
|
将样地逐个植株胸径、株高带入上述模型估算出各株杉木的地上生物量,再利用地上地下生物量比推算地下生物量。同理, 杉木优势林和含杉木的混交林中的马尾松和其他常绿阔叶林生物量也采用本研究组的其他模型研究结果(刘其霞等,2005),估算出各株乔木的生物量逐株累加求得各个公益林样地乔木层生物量。在样方外围选取不同树种、等级的下木层标准木3~5株,分别将各树种标准木的干、枝、叶、果和根取样并带回实验室,分别测鲜质量和干质量,根据各树种各器官含水率求出各树种标准木各器官的干质量,各树种标准木各器官干质量相加可得各树种标准木生物量,按株数推算各树种各单位面积生物量,各树种单位面积生物量相加即得下木层单位面积生物量。在草本层小样方外围采用整株挖掘法取草样、称鲜质量,并带回实验室烘干、称干质量,计算含水率,通过换算求得各单位面积的生物量。
乔木层、下木层和草本层各层生物量之和即为整个群落的生物量。
2.3 数据分析参照浙江林业自然资源的林龄划分标准(刘安兴等, 2002)和对生态公益林的要求,将浙江省杉木生态公益林划分为3个年龄级:10年以下为幼龄林,11~20年为中龄林,20年以上为成熟林(刘国华等,2000; 黄承才等,2006)。
根据杉木占乔木层的相对密度以及相对重要值(金则新,2001),将杉木相对密度在70 %以上,相对重要值在0.60以上的样地划为杉木优势林地,其余为含杉木的混交林地(黄承才等,2006)。这些样地中包括杉木优势林样地28个,含杉木的混交林样地39个。
不同森林植被因其群落组成、年龄结构、林分起源的差异,其碳储量转换率略有不同,杉木生态公益林植被碳储量基于其生物量乘以转换比率(也称碳素密度)。周玉荣等(2000)和王效科等(2001)采用国际上常用的转换率0.45;刘国华等(2000)采用0.5;方晰等(2002)研究得出杉木树叶、树枝、树根、树干、树皮和球果的碳素密度分别为0.491 6,0.460 5,0.472 4,0.474 4,0.500 3和0.469 9。本研究中杉木林的碳素密度取以上加权平均值,即为0.48;含杉木的混交林常绿阔叶树种较多,故采用目前文献中的碳密度最大值0.5(Fang et al., 2001; 刘国华等,2000)。
3 结果与分析 3.1 各龄级杉木生态公益林乔木层生物量与林分密度幼龄、中龄和成熟杉木生态公益林乔木层的平均生物量分别为43.82,54.99和64.40 t·hm-2,其中中龄林比幼龄林增长了25.5 %,成熟林比中龄林增长了17.1 %(表 2)。而这3个阶段的杉木林平均密度无显著差异,密度因素的影响在此可以忽略。
|
|

杉木优势林乔木层平均生物量在前2个年龄级保持在48 t·hm-2左右,其密度变化也不大(表 3)。说明10年以后杉木优势林整个林分的生物量增长缓慢,对碳积累的贡献较小。在林业经营过程中,通常对15年左右林龄的杉木进行人工间伐,使得密度下降,再经过10余年的生长,乔木层生物量略有增长(增幅 < 9 %)。
|
|
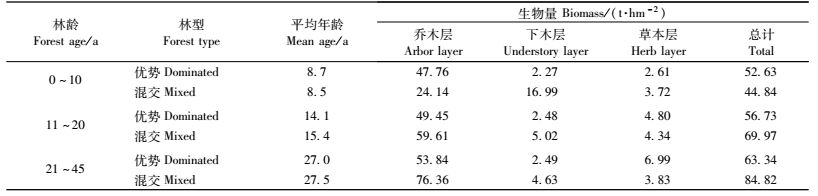
含杉木混交林的乔木层平均生物量随着林龄增长明显增长:幼龄林为24.14 t·hm-2; 中龄林为59.61 t·hm-2(表 3),比幼龄林增长了147%;成熟林为76.36 t·hm2,比中龄林增长了28.1%。
3.2 杉木生态公益林群落生物量分配杉木优势林的乔木层所占比例逐渐降低,中龄和成熟阶段比同年龄级的含杉木混交林的低。杉木优势林下木层生物量(2.27~2.49 t·hm-2)及其占总生物量的比例变化较小(4.31%~4.94 %),且都低于含杉木的混交林。杉木优势林草本层生物量在幼龄时低于含杉木混交林,至中龄时比例增加,直到超过后者(表 3)。
含杉木混交林的群落生物量随着林龄增长也很快:幼龄林为44.84 t· hm-2,中龄林为69.97 t· hm-2,成熟林为84.82 t·hm-2(表 3)。除了含杉木混交林的幼龄林外,乔木层生物量所占比例都在85 %以上。含杉木混交林的幼龄林乔木层郁闭度小,下层有一定比例的灌木和草本生长,因此下木层生物量(16.99 t·hm-2)所占比例达37.88 %,以后逐渐降低。
3.3 杉木生态公益林群落碳储量幼龄、中龄和成熟杉木生态公益林乔木层碳储量分别为20.82,26.12和32.97 t·hm-2,各年龄级间年增量为0.53~0.69 t·hm-2,增幅约为25 %。杉木优势林各年龄间年增量为0.08~0.21 t·hm-2,含杉木的混交林各年龄间年增量为0.84~1.77 t·hm-2(表 4)。
|
|
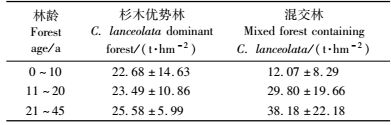
浙江生态公益林中的10年以下杉木优势林乔木层生物量和按照商用林方式管理的杉木林差别不显著(等方差t检验P=0.344 9),10年后差别越来越大(异方差t检验P < 0.001),以公益林方式管理的杉木优势林的生物量10年以后生物量显著低于文献中的杉木林生物量(秦建华等,1996; 俞新妥等,1997; 陈楚莹等,2000; 方奇,2000; 田大伦等,2003),而文献中杉木林大部分还是按照商用林方式进行管理(陈楚莹等,2000; Zhang et al., 2007)。从生态效益的碳积累角度出发必须进行改造,如果以人工方式间伐补种改造成含杉木的混交林或常绿阔叶林,生物量还会随着林龄增长继续增长(刘其霞等,2005)。另一方面,大部分文献对生物量的估计一般是基于典型的试验样地,统计结果可能偏高; 本研究是对生态公益林现状的分析,样本大且跨越全省,本研究的杉木林生物量平均值对浙江森林而言具有一定的代表性。
杉木优势林依靠人工管理和高密度种植,在前10年的幼龄阶段就达到较高的乔木层生物量; 但在中龄林及成熟林阶段杉木优势林乔木层生物量增加很少,都低于同龄级的含杉木混交林,对碳积累贡献很小; 必须循序渐进改造为含杉木的混交林,并逐渐向针阔混交林过渡(沈琪等,2005),直到演替为碳积累潜力最大的常绿阔叶林(Zhang et al., 2007)。
浙江省碳储量自20世纪50年代到90年代初一直下降,1993年以后才开始上升; 1994—1998年浙江省碳储量为19.9 t·hm-2,不到全国平均水平44.9 t·hm-2的一半(方精云等,2001)。2001年浙江省总体碳贮量低只占全国的0.74%(王效科等,2001)。本研究发现杉木生态公益林碳年增量约为0.53 t·hm-2,虽然高于全国森林平均水平0.24 t·hm-2(刘国华等,2000),但是杉木纯林或优势林初期是依靠高密度种植和人工管理,才有高的生物量,随着生态公益林的建设,封山育林后对其干扰减少,碳储量年增量很小(表 4)。若逐步改造为碳储量年增量较高的含杉木的混交林,杉木生态公益林的碳积累量还会进一步提高:若杉木林改造为含杉木的混交林,碳储量每年至少增加0.84 t·hm-2; 若不改造,每年至多增加0.21 t·hm-2。
俞新妥等(1997)和陈楚莹等(2000)认为杉木老龄林必然要被亚热带阔叶树种所代替,因为耐荫的常绿或落叶阔叶植物可逐渐在杉木群落内定居下来,成为杉木群落的组成成分。本研究中按经济林管理方式间伐的杉木林经过10余年的生长,乔木层基本没有其他树种成长起来,而下木层只有少量常绿灌木种类。这说明常绿或落叶树种侵入杉木优势林还有一定困难。要更快降低杉木在林中的比例,可增加间伐强度适当降低密度,宜采用短轮伐期作业(叶镜中等,1983),甚至可以通过人工管理,择伐后补种苦槠、青冈(Cyclobalanopsis sp.)、木荷、檵木(Loropetalum sp.)、柃木(Eura japonica)和冬青(Ilex sp.)等其他常绿树种来加速恢复过程,从而迅速恢复亚热带地区森林生态系统功能。
陈楚莹, 廖利平, 汪思龙. 2000. 杉木人工林生态学[M]. 北京: 科学出版社.
|
方奇. 2000. 不同密度杉木幼林系统生产力和生态效益研究[J]. 林业科学, 36(1): 28-35. DOI:10.11707/j.1001-7488.20000105 |
方精云, 陈安平. 2001. 中国森林植被碳库的动态变化及其意义[J]. 植物学报, 43(9): 967-973. |
方精云, 刘国华, 朱彪, 等. 2006. 北京东灵山三种温带森林生态系统的碳循环[J]. 中国科学D辑:地球科学, 36(6): 533-543. |
方精云, 沈泽昊, 唐志尧, 等. 2004. "中国山地植物物种多样性调查计划"及若干技术规范[J]. 生物多样性, 12(1): 5-9. |
方晰, 田大伦, 项文化. 2002. 速生阶段杉木人工林碳素密度、贮量和分布[J]. 林业科学, 38(3): 14-19. DOI:10.11707/j.1001-7488.20020303 |
冯宗炜, 陈楚莹, 张家武. 1982. 湖南会同地区马尾松林生物量的测定[J]. 林业科学, 18(2): 127-134. |
黄承才, 张骏, 江波, 等. 2006. 浙江省杉木生态公益林凋落物及其与植物多样性的关系[J]. 林业科学, 42(6): 7-12. |
胡会峰, 王志恒, 刘国华, 等. 2006. 中国主要灌丛植被碳储量[J]. 植物生态学报, 30(4): 539-554. |
金则新. 2001. 浙江天台山落叶阔叶林优势种群结构与动态分析[J]. 浙江林学院学报, 183(3): 245-251. |
林生明, 徐土根, 周国模. 1991. 杉木人工林生物量的研究[J]. 浙江林学院学报, 8(3): 288-294. |
刘安兴, 张正寿, 丁衣冬. 2002. 浙江林业自然资源:森林卷[M]. 北京: 中国农业科学技术出版社.
|
刘国华, 傅伯杰, 方精云. 2000. 中国森林碳动态及其对全球碳平衡的贡献[J]. 生态学报, 20(5): 733-740. |
刘其霞, 常杰, 江波, 等. 2005. 浙江省常绿阔叶生态公益林生物量[J]. 生态学报, 25(9): 2139-2144. |
秦建华, 姜志林. 1996. 杉木林生物量及其分配变化的规律[J]. 生态学杂志, 15(1): 1-7. |
田大伦, 康文星, 文仕知, 等. 2003. 杉木林生态系统学[M]. 北京: 科学出版社.
|
沈琪, 张骏, 朱锦茹, 等. 2005. 浙江省生态公益林植被恢复过程中物种组成和多样性的变化[J]. 生态学报, 25(9): 2131-2138. |
王效科, 冯宗炜, 欧阳志云. 2001. 中国森林生态系统的植物生物量和碳密度研究[J]. 应用生态学报, 12(1): 13-16. |
叶镜中, 姜志林. 1983. 苏南丘陵杉木人工林的生物量结构[J]. 生态学报, 3(1): 7-14. |
俞新妥, 范少辉, 林思祖, 等. 1997. 杉木栽培学[M]. 福州: 福建科学技术出版社.
|
浙江森林编辑委员会. 1993. 浙江森林[M]. 北京: 中国林业出版社.
|
周国逸, 闫俊华. 2000. 生态公益林补偿理论与实践[M]. 北京: 气象出版社.
|
周玉荣, 于振良, 赵士洞. 2000. 我国主要森林生态系统碳贮量和碳平衡[J]. 植物生态学报, 24(5): 518-522. |
Brown S L, Schrode P, Kern J S. 1999. Spatial distribution of biomass in forests of the eastern USA[J]. Forest Ecology and Management, 123: 81-90. DOI:10.1016/S0378-1127(99)00017-1 |
Cairns M A, Olmsted I, Granados J, et al. 2003. Composition and aboveground tree biomass of a dry semi-evergreen forest on Mexico's Yucatan Peninsula[J]. Forest Ecology and Management, 186: 125-132. DOI:10.1016/S0378-1127(03)00229-9 |
Fang J Y, Chen A P, Peng C H, et al. 2001. Changes in forest biomass carbon storage in China between 1949 and 1998[J]. Science, 292: 2320-2323. DOI:10.1126/science.1058629 |
Zhang J, Ge Y, Chang J, et al. 2007. Carbon storage by ecological service forests in Zhejiang Province, subtropical China[J]. Forest Ecology and Management, 245: 64-75. DOI:10.1016/j.foreco.2007.03.042 |