 2010, Vol. 46
2010, Vol. 46文章信息
- 文仕知, 田大伦, 杨丽丽, 方晰
- Wen Shizhi, Tian Dalun, Yang Lili, Fang Xi
- 桤木人工林的碳密度、碳库及碳吸存特征
- Carbon Density, Carbon Stock and Carbon Sequestration in Alnus cremastogyne Plantation
- 林业科学, 2010, 46(6): 15-21.
- Scientia Silvae Sinicae, 2010, 46(6): 15-21.
-
文章历史
- 收稿日期:2009-04-17
- 修回日期:2009-07-26
-
作者相关文章


2. 南方林业生态应用技术国家工程实验室 长沙 410004
2. State Key Laboratory of Ecological Applied Technology in Forest Area of South China Changsha 410004
当前,大气中CO2等温室气体的浓度正在逐年增高,全球气候变暖对人类的影响越来越显著,已成为国际社会最关注的全球性环境问题,对CO2 吸收、固定和排放的研究也已成为全球变化研究的热点和前沿(Fang et al., 2001; Caldeira et al., 2000; 方精云等,2007)。据实测,当前地表和大气温度的上升,70%~80%是由于大气中CO2的增加造成的(李克让等,1996)。森林是陆地生态系统的主体,在调节全球碳收支平衡、减缓大气CO2等温室气体浓度上升等方面具有不可替代的作用。方精云(2001;2007)、王效科等(2000;2001)、刘国华等(2000)和周玉荣等(2000)分别利用我国森林资源调查资料,结合我国森林生态系统生物量和生产力研究的基础上,推算我国近50年来森林植被的碳库及其动态,为评价北半球中高纬度地区碳库和我国森林碳汇功能做出了贡献。近十几年来,随着绿化工程和退耕还林工程的实施,我国人工林面积在森林总面积中的比重快速增加。人工林在CO2的吸收和固定及减缓全球气候变化等方面的作用也随之得到重视(Winjum et al., 1997)。目前,人工林碳贮量与碳平衡的研究国内外已有较多报道(Laclau,2003; Specht et al.,2003;方晰等,2002; 何宗明等,2003; 雷丕锋等,2004; 包维楷等,2005; 李轩然等,2006; 马明东等,2008)。
四川桤木(Alnus cremastogyne)属桦木科(Betulaceae)桤木属,为中国特有种,是喜温、喜湿、耐水和耐瘠薄的优良纤维材和水土保持树种,是适合湖南退耕还林工程和生态建设工程的优良树种。截止2005年湖南省桤木栽培面积已超过8.3万hm2,根据当前的发展形势,桤木造林仍是湖南今后的造林重点,年造林面积在现有基础上还有较大增长空间。与我国其他类型的人工林相比,桤木是非豆科结瘤树种,具有较强的固氮能力(邓廷秀等,1987),能够增加土壤氮素含量,对植物群落的生长及土壤有机碳含量有显著影响(Vogel et al., 1998)。目前有关桤木的研究主要集中在桤木改良土壤效应、群落特征及生物量分析、桤木光合生理特性研究、扦插繁殖技术、氮素积累、生物产量与生物循环等(刘国凡等,1985; 向永国等,1995; 王军辉等,2000; 刘贤词等,2007; 杨丽丽等,2008),有关桤木人工林碳固定能力的研究未见报道。本研究通过分析不同年龄桤木人工林生态系统的碳密度、碳库空间分布及碳吸存变化和估算湖南省现有桤木林植被层的碳库和碳吸存潜力,为在区域或国家尺度上估算森林生态系统碳库以及碳平衡的估算提供基础数据,为评价桤木人工林的生态服务功能提供科学依据。
1 试验地概况试验地设置于湖南湘西花垣县,地理坐标为109°15′—109°38′ E,28°10′—28°38′ N,地处云贵高原东缘、武陵山脉南段中部地区,多为山地地貌。本区气候属亚热带季风气候,四季分明,平均气温16 ℃,无霜期约270天,年均降雨量1 420.9 mm,日照时数1 324.5 h,海拔从212~1 197 m,相对高度达985 m,成土母岩以石灰岩为主,土壤主要为红壤和山地黄壤; 土壤呈微酸性至中性,地带性植被为常绿阔叶林。试验地林分特征如表 1所示。
|
|
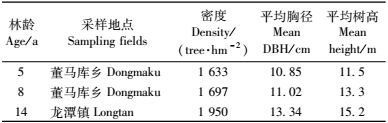
在花垣县桤木人工林内选择5, 8, 14年生3个年龄段的桤木林固定样地各2块,样地面积均为30 m×20 m,并进行每木调查。根据胸径、树高和冠幅等测树因子,在每样地选取3株平均木,即每年龄段共选6株平均样木并伐倒,按2 m区分段,测定各区分段的干、皮、枝、叶各器官的鲜质量,同时按树高不同层次分组分别采集伐倒木的分析样品,带回室内采用烘干法测定其含水量。地下部分采用“分层挖掘法”(0~15,15~30,>30 cm)分别测定根头、大根(>0.5 cm)、粗根(0.2~0.5 cm)、细根(≤0.2 cm)鲜质量同时采集分析样品。分析样品在80 ℃烘干至恒量,计算出各器官干质量。因桤木人工林各径阶分化不明显,故用平均木法对全林生物量进行估测,考虑到同龄人工林特点,采用年平均生物量作为净生产力的估算指标(潘维俦等,1981),由于桤木是落叶乔木树种,树叶的生物量是当年新萌发出来的,因此桤木人工林树叶净生产力就是当年的生物量。
2.2 林下植被生物量在每个固定样地的对角线上离4个角各1 m处和样地中心设1 m×1 m小样方5个,记录每个小样方内的植物种类,分地上部分和地下部分,采用“全挖实测法”分别测定其鲜质量,同种植物的相同器官取混合样品,死地被物全部测定生物量,取混合样品,80 ℃烘干至恒量后再估算干质量,同时用作分析样品。
2.3 土壤样品采集在各个样地内,按0~15,15~30,30~60 cm 3个层次随机采集3~4个样点的土壤分析样品,同时测定土壤密度,根据土壤密度计算各土层单位面积土壤质量。
2.4 分析样品中碳含量的测定、碳库及碳吸存量的计算方法土壤样品置于室内风干,然后磨碎,再以105 ℃烘干至恒量。植物、土壤样品中的碳密度采用重铬酸钾-水合加热法测定(田大伦,2004)。数据统计分析采用SPSS 13.0软件包进行。用各器官的碳含量与其生物量来估算林分碳库量; 用各器官的碳含量与其生物量的年增量相乘来估算林分的碳吸存量; 土壤碳库采用单位面积土层质量与相应土层碳含量的乘积。
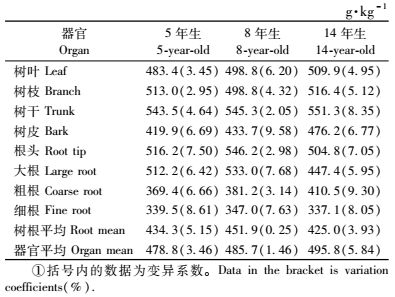
3 结果与分析 3.1 不同年龄桤木人工林生态系统中各组分碳密度的动态 3.1.1 桤木不同器官碳密度如表 2所示,5年生桤木各器官碳密度变化范围为339.5~543.5 g·kg-1,算术平均值为478.8 g·kg-1,8年生的为347.0~546.2 g·kg-1,算术平均值为485.7 g·kg-1,14年生的为337.1~551.3 g·kg-1,算术平均值为495.8 g·kg-1,变异系数在0.25%~9.58%之间,随桤木林年龄的增长,各器官碳密度的变化规律不明显; 不同器官按碳密度高低排列大致为树干>树枝>树叶>树根>树皮。树根中,均以根头的碳密度最大,粗根次之,细根最小,变异系数在0.25%~9.58%。差异显著性分析表明,不同年龄之间的同器官碳密度平均值的差异均未达到显著水平(P>0.05)。
|
|
如表 3所示,不同年龄桤木林下地被物的碳密度均表现为:活地被物>死地被物,活地被物层中表现为地上部分高于地下部分。5年生桤木林中,活地被物与死地被物的平均碳密度分别为410.1和391.3 g·kg-1,8年生的分别为397.0和332.4 g·kg-1,14年生的分别为434.7和318.6 g·kg-1,林下地被物各器官碳密度变异系数为2.90%~13.03%。随着桤木林年龄的增长,林下地被物的碳密度变化规律也不明显。差异显著性分析表明:不同年龄之间桤木林下活地被物层、死地被物层的差异同样也未达到显著水平(P>0.05)。
|
|
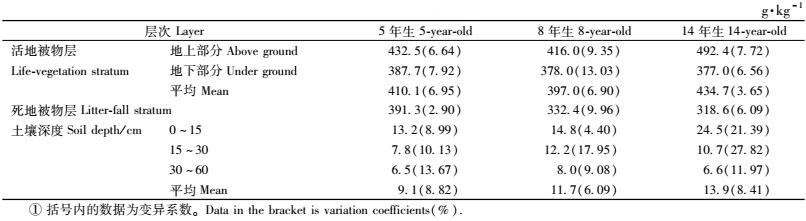
从表 3可以看出,桤木林地土壤层(0~60 cm)碳密度平均值随着林龄的增长逐渐增高,从5年生的9.1 g·kg-1增加到14年生13.9 g·kg-1。不同年龄桤木林地土壤碳密度均随着土层深度的增加呈现出递减趋势。方差分析结果表明,不同年龄之间同一土层碳密度的差异均未达到显著水平(P>0.05)。
3.2 桤木林各器官的碳贮量与分配由表 4可以看出,桤木林的生物量和碳贮量均随着林龄的增长而增加,5年生到14年生,生物量由51.23 t·hm-2增加到95.31 t·hm-2,碳贮量由25.88 t·hm-2增加到49.63 t·hm-2,尤以树干增加最为明显,树干碳贮量占林分碳贮量的百分比从5年生的58.85%增加到14年生的59.74%。各器官中的碳贮量基本上与其生物量成正比例关系,树干的碳贮量最高,如5年生时,树干生物量占林分生物量的54.52%,其碳贮量所占比例为58.85%,14年生时占56.44%,其碳贮量所占的比例更高,为59.74%。地上部分的碳贮量占桤木林碳贮量的84.98%以上。
|
|
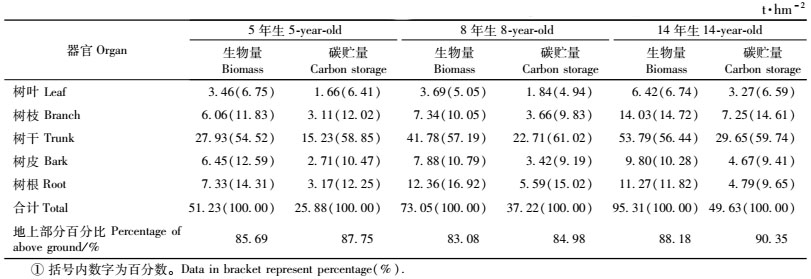
表 5表明,5,8和14年生桤木人工林土壤(0~60 cm)层的碳储量分别为68.61,83.55和77.69 t·hm-2,且呈现出明显的垂直分布,即土壤碳储量随着土壤层深度的增加而逐渐下降。5,8和14年生桤木林地土壤表层(0~15 cm)的碳储量分别占土壤(0~60 cm)层碳储量的38.71%,34.63%和46.87%,而0~30 cm土壤层中的碳储量分别占61.72%,61.68%和66.55%。可见,林地土壤0~30 cm土层保存了绝大部分的碳。
|
|
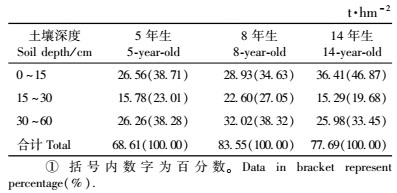
如表 6所示,5,8和14年生桤木林生态系统中的碳库分别为95.89,122.12和130.75 t·hm-2,主要由植被层、死地被物层和土壤层组成,按其碳库的大小顺序排列为:土壤层>植被层>死地被物层。植被层的碳库随林分年龄的增长而增加,5,8和14年生桤木林生态系统中,植被层的碳库分别为26.63,37.76和51.54 t·hm-2,分别占整个生态系统碳库总量的27.77%,30.18%和39.42%。其中乔木层碳贮量随林分年龄的增长而增加的程度最为明显,5年生时,乔木层的碳贮量占植被层的97.18%,占整个生态系统的26.99%,8年生时分别占98.52%和29.73%,14年生时分别占96.29%和37.96%。5,8和14年生桤木人工林林下植被碳贮量分别占植被层碳贮量的2.82%,1.48%和3.71%。由此可见,在桤木人工林生态系统中,植被层的碳贮量主要取决于乔木层的碳贮量。林下死地被物层的碳贮量随着林龄的增长而增加,5年生时为0.65 t·hm-2,14年生时增加到1.52 t·hm-2,分别占整个生态系统碳贮量的0.68%,1.16%。林地土壤层(0~60 cm)的碳贮量相当可观,在不同年龄的桤木林中均在68.61 t·hm-2以上,占整个生态系统碳贮量的59.42%以上。
|
|
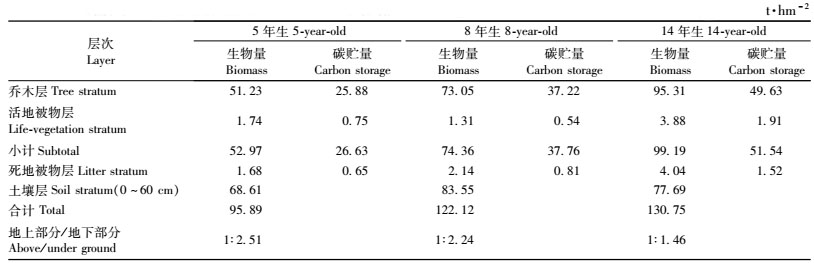
从表 6还可以看出,桤木人工林生态系统中地上部分与地下部分碳贮量之比为:5年生为1:2.51,8年生为1:2.24,14年生为1:1.46,随着林分年龄的增长,地上部分植物层的生物量明显增加,其碳贮量也随着增加,而土壤中的碳储量增加速度缓慢。因此,随着林分年龄的增加,地上部分与地下部分碳贮量之比逐渐减小。
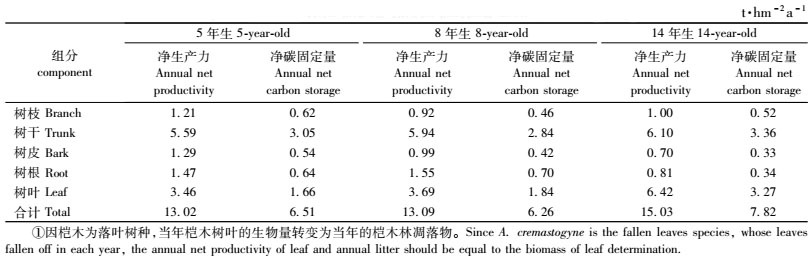
3.5 桤木林净第一性生产力与年净固定碳量的初步估算由表 7可以看出,5,8和14年生桤木人工林生态系统的净第一性生产力分别为13.02,13.09和15.03 t·hm-2a-1,其年净固定碳量分别6.51,6.26和7.82 t·hm-2a-1,生物量当年净增量(除落叶外)分别为9.56,9.40和8.61 t·hm-2a-1,净积累碳量分别是4.85,4.42和4.55 t·hm-2a-1,分别占桤木林当年净固定碳量的74.50%,70.61%和58.18%,均以树干的年固定碳量为最高。当年凋落物中固定的碳量分别为1.66,1.84和3.27 t·hm-2a-1,分别占当年净固定碳素量的25.50%,29.39%和41.82%。
|
|
桤木各器官碳密度的算术平均值随着林龄的增长而增高,5,8和14年生分别为478.8,485.7和495.8 g·kg-1,桤木各器官碳密度大小排序大致为:树干>树枝>树叶>树根>树皮。各器官碳密度随着林木年龄的增长,变化规律不明显,且不同年龄之间,各器官碳密度的差异均不显著。与此相比,10,11和14年生杉木(Cunninghamia lanceolata)林各器官碳密度算术平均值分别为475.6,477.9和480.7 g·kg-1(田大伦等,2004),18年生樟树(Cinnamomum camphora)人工林的为498.6 g·kg-1(雷丕锋等,2004),32年生楠木林(Phoebe bourmei)的为493.1 g·kg-1(马明东等,2008),40年生栓皮栎(Quercus variabilis)的叶、枝、干、皮和根的碳密度分别为521.0,486.0,512.0,486.0和452.0 g·kg-1(阮宏华等,1997); 而海南岛尖峰岭热带山地雨林中主要树种的叶、枝、干、皮和根的碳密度分别为458.4,465.3,579.0,456.2和539.0 g·kg-1,不同树种各组分内碳密度变化不大,其样本变异系数为6.5%~15%(李意德等,1998)。显然,同一地区的不同树种或不同地区不同树种间各器官碳密度有一定的差异,但它们的变化大都在450.0~550.0 g·kg-1范围之内。
随着桤木林年龄的增长,林下地被物的碳密度变化规律也不明显。这可能是由于草本层植物种类较多所决定的。乔木层中地上部分桤木各器官碳密度平均值明显高于林下活地被物地上部分的碳密度,这可能与林下植物种类、林下的环境条件有关。同一林分中乔木层平均碳密度高于活地被物层。土壤(0~60 cm)层平均碳密度随着桤木林年龄的增长逐渐增加,且均呈现出明显的垂直分布。
桤木林的碳贮量随着年龄的增长而增加,从5年生25.88 t·hm-2增加到14年生的49.63 t·hm-2,且以树干最为明显,树干碳贮量占林分碳贮量的百分比从5年生的58.85%增加到14年生的59.74%,高于湖南会同10,11和14年生杉木林,18年生樟树林树干碳贮量所占林分碳贮量的百分比(方晰等,2002; 方晰,2004; 雷丕锋等,2004)。5,8和14年生桤木林地上部分碳贮量分别占桤木林分碳贮量的87.75%,84.44%和90.35%。据报道,湖南会同10,11和14年生杉木人工林地上部分碳贮量分别占杉木林分碳贮量的85.35%,84.73%和86.54 %(方晰等,2002; 方晰,2004),福建三明33年生的福建柏(Fokienia hodginsii)和杉木人工林地上部分碳贮量分别占林分碳贮量的85.71%和81.13%(何宗明等,2003)。
5,8和14年生桤木人工林土壤层(0~60 cm)的碳素含量分别为68.61,83.55和77.69 t·hm-2,低于世界土壤平均碳贮量(189.00 t·hm-2)和我国森林土壤平均碳贮量(193.55 t·hm-2)(周玉荣等,2000),也低于湖南会同杉木人工林土壤碳贮量(14年生108.20 t·hm-2)(方晰等,2002; 方晰,2004),高于江苏南部丘陵地区的次生栎林(secondary oak forests)土壤(0~80 cm)的碳贮量(69.70 t·hm-2)(阮宏华等,1997),究其原因可能是采集土壤样品时,分层的差异,或者是在估算土壤碳储量时所测算的土壤深度不同。由于森林动植物的残体和枯枝落叶作为土壤有机碳的主要来源,并在气候、生物等因素的作用下,在林地土壤中形成了层次结构,其碳储量也将随着土壤深度的不同而发生变化。桤木人工林林地土壤的碳储量随着土壤深度的增加而减少,其中0~30 cm的土壤层中的碳贮量分别占其总碳贮量的61.72%,61.68%和66.55%。也正由于土壤中的碳主要分布在0~30 cm土层,以及人类各种经营活动也主要发生在0~30 cm土层。因此,人类经营活动方式对土壤中的碳也就产生巨大的影响,这也往往决定了森林土壤中的碳库是“碳源”或是“碳汇”的作用。
不同年龄阶段桤木人工林生态系统地上部分和地下部分碳贮量之比为:5年生为1:2.51,8年生为1:2.24,14年生为1:1.46。随着桤木林龄的增长,地上部分与地下部分碳贮量之比有逐渐下降的趋势。根据Dixon(1994)对全球森林生态系统碳库贮量和碳通量的研究可知,全球范围内植被与土壤碳贮量之比平均为0.46,全球低纬度地区(0~25°)为0.83~1.25,亚洲低纬度地区该比值为0.95~1.25,我国(按照其文章中划分属于中纬度地区)25°~50°平均为0.84。周玉荣等(2000)对我国主要森林生态系统碳贮量的研究表明,我国森林植被与土壤碳储量的比值为0.19~0.95,平均为0.36。可见,无论是在全球范围还是在全国范围内,土壤碳储量是植被的2倍左右。
5,8和14年生桤木人工林生态系统碳库分别为:95.89,120.82和130.75 t·hm-2,明显低于我国森林生态系统平均碳贮量(258.83 t·hm-2)(周玉荣等,2000)。桤木人工林生态系统的碳库主要由植被层、死地被物层和土壤层组成的,按其碳贮量大小顺序排列为:土壤层>植被层>死地被物层。由此可见,森林生态系统中的林地土壤层和植被层是碳的一个极重要的贮存库,表明了保护好现有的森林植被对维持陆地生态系统的碳贮量有着重要的意义。死地被物层的碳贮量虽然远小于土壤层和植被层的碳贮量,但它是森林涵养水源的主要功能层,也是森林土壤碳的主要来源,对森林生态系统的碳循环起着极为重要作用。
森林生态系统生产力研究的主要内容之一是要确定系统同化CO2的能力。5,8和14年生桤木林年净固定碳量分别6.51,6.26和7.72 t·hm-2a-1,生物量当年净增碳量分别为4.85,4.42和4.55 t·hm-2a-1。据报道,海南尖峰岭热带雨林年净固定碳量为8.62 t·hm-2a-1,生物量当年碳同化净增量为3.82 t·hm-2a-1(李意德等,1998); 江苏南部丘陵地区国外松林(Loblolly pine)生物量的碳同化净增量为5.79 t·hm-2a-1(阮宏华等,1997); 福建三明33年生的福建柏和杉木人工林年净固定碳量分别9.907和6.450 t·hm-2a-1(何宗明等,2003)。
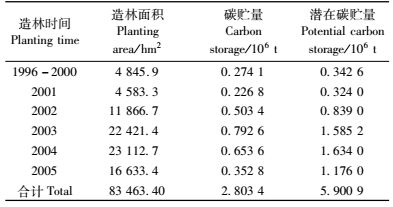
四川桤木人工林数量成熟年龄为9~10年(代顺民等,2007)。本研究中将10年作为桤木的成熟年龄,根据湖南省现有桤木林分布面积(徐清乾等,2006),并以本研究的5,8和14年生桤木人工林的净第一性生产量及其固定碳量的平均值(即净生产力为13.71 t·hm-2a-1,碳固定量为6.86 t·hm-2a-1)估算湖南省桤木林现有碳库和潜在碳库。如表 9所示,湖南省现有桤木林植被的碳库为2.803 4×106t,占湖南省森林植被的碳库的7.24%(王效科等,2001),其潜在碳库为5.900 9×106 t,碳吸存潜力为3.097 6×106t。如果以1 t碳等于3.67 t CO2计(王效科等,2000),则折合成湖南现有桤木林植被CO2吸存潜力为11.368 0×106t,即按桤木轮伐期10年计,现有的桤木林还能吸收11.368 0×106t CO2。湖南省桤木林现有碳库为潜在碳库的47.51%,即现有的桤木林生态系统的实际碳贮量为潜在的一半左右,说明湖南省桤木林林分质量处于中等水平。
|
|
包维楷, 雷波, 冷俐. 2005. 六种人工针叶幼林下地表苔藓植物生物量与碳贮量[J]. 应用生态学报, 16(10): 1817-1821. DOI:10.3321/j.issn:1001-9332.2005.10.004 |
代顺民, 唐巍, 唐大云. 2007. 四川桤木短轮伐期用材林合理采伐年龄研究[J]. 四川林勘设计, 6(2): 18-20. |
邓廷秀, 刘国凡. 1987. 桤柏混交林的初步研究[J]. 植物生态学与地植物学学报, 11(1): 59-67. |
方精云, 陈安平. 2001. 中国森林植被碳库的动态变化及其意义[J]. 植物学报, 43(9): 967-973. |
方精云, 郭兆迪, 朴世龙, 等. 2007. 1981—2000年中国陆地植被碳汇的估算[J]. 中国科学D辑:地球科学, 37(6): 804-812. |
方晰, 田大伦, 项文化. 2002. 速生阶段杉木人工林C密度、贮量和分布[J]. 林业科学, 38(3): 14-19. DOI:10.11707/j.1001-7488.20020303 |
方晰. 2004. 杉木人工林生态系统碳贮量与碳平衡的研究. 中南林业科技大学博士学位论文.
|
何宗明, 李丽红, 王义祥, 等. 2003. 33年生福建柏人工林碳库与碳吸存[J]. 山地学报, 21(3): 298-303. |
雷丕锋, 项文化, 田大伦, 等. 2004. 樟树人工林生态系统碳素贮量与分布研究[J]. 生态学杂志, 23(4): 25-30. |
李克让, 陈育峰, 刘世荣, 等. 1996. 减缓及适应全球气候变化的中国林业对策[J]. 地理学报, 5(增): 109-119. |
李轩然, 刘琪, 陈永瑞, 等. 2006. 千烟洲人工林主要树种地上生物量的估算[J]. 应用生态学报, 17(8): 1382-1388. |
李意德, 吴仲民, 曾庆波, 等. 1998. 尖峰岭热带山地雨林群落生产和二氧化碳同化净增量的初步研究[J]. 植物生态学报, 22(2): 127-134. |
刘国凡, 邓廷秀. 1985. 土壤条件与桤木结瘤固氮的关系[J]. 土壤学报, 22(3): 251-257. |
刘国华, 傅伯杰, 方精云. 2000. 中国森林碳动态及其对全球碳平衡的贡献[J]. 生态学报, 20(5): 733-740. |
刘贤词, 文仕知, 冯汉华, 等. 2007. 四川桤木人工林不同年龄段生物量的研究[J]. 中南林业科技大学学报, 27(2): 83-86. |
马明东, 江洪, 刘跃建. 2008. 楠木人工林生态系统生物量、碳含量、碳贮量及其分布[J]. 林业科学, 44(3): 34-39. DOI:10.11707/j.1001-7488.20080310 |
潘维俦, 田大伦. 1981. 森林生态系统第一性生产量的测定技术与方法[J]. 湖南林业科学, (2): 1-12. |
阮宏华, 姜志林, 高苏铭. 1997. 苏南丘陵主要森林类型碳循环研究:含量与分布规律[J]. 生态学杂志, 16(6): 17-21. |
田大伦, 方晰, 项文化. 2004. 湖南会同杉木人工林生态系统的碳素密度[J]. 生态学报, 24(11): 2382-2386. DOI:10.3321/j.issn:1000-0933.2004.11.006 |
田大伦. 2004. 杉木林生态系统定位研究方法[M]. 北京: 科学出版社: 320-341.
|
王军辉, 顾万春, 李斌, 等. 2000. 桤木优良种源/家系的选择研究:生长的适应性和遗传稳定性分析[J]. 林业科学, 36(3): 59-66. DOI:10.11707/j.1001-7488.20000311 |
王效科, 冯宗炜. 2000. 中国森林生态系统中植物固定大气碳的潜力[J]. 生态学杂志, 19(4): 72-74. |
王效科, 冯宗炜. 2001. 中国森林生态系统的植物碳储量和碳密度研究[J]. 应用生态学报, 12(1): 13-16. |
向永国, 王金锡. 1995. 桤柏混交林能量代谢中能量吸收、固定、积累和损耗研究[J]. 四川林业科技, 16(1): 1-11. |
徐清乾, 许忠坤, 蒋玉璋. 2006. 四川桤木引种湖南生长状况及适应性研究[J]. 湖南林业科技, 33(6): 20-24. |
杨丽丽, 文仕知, 王珍珍. 2008. 不同立地条件下桤木人工林生物量和生产力的比较[J]. 中南林业科技大学学报, 28(1): 122-126. |
周玉荣, 于枕良, 赵士洞. 2000. 我国主要森林生态系统碳贮量和碳平衡[J]. 植物生态学报, 24(5): 518-522. |
Caldeira K, Duffy P B. 2000. The role of the southern ocean in up take and storage of anthropogenic carbon dioxide[J]. Science, 287: 620-622. DOI:10.1126/science.287.5453.620 |
Dixon R K. 1994. Carbon pools and flux of global forest ecosystems[J]. Science, 263: 185-189. DOI:10.1126/science.263.5144.185 |
Fang J Y, Chen A P, Peng C H, et al. 2001. Changes in forest biomass carbon storage in China between 1949 and 1998[J]. Science, 292: 2320-2322. DOI:10.1126/science.1058629 |
Laclau P. 2003. Biomass and carbon sequestration of ponderosa pine plantations and native cypress forests in northwest Patagonia[J]. Forest Ecology and Management, 180: 317-333. DOI:10.1016/S0378-1127(02)00580-7 |
Specht A, West P W. 2003. Estimation of biomass and sequestered carbon on farm forest plantations in northern New South Wales, Australia[J]. Biomass and Bioenergy, 25: 363-379. DOI:10.1016/S0961-9534(03)00050-3 |
Vogel J G, Gower S T. 1998. Carbon and nitrogen dynamics of boreal jack pine stands with and without a green alder understory[J]. Ecosystems, 1: 386-400. DOI:10.1007/s100219900032 |
Winjum J K, Schroeder P E. 1997. Forest plantations of the world: Their extent, ecological attributes, and carbon storage[J]. Agricultural and Forest Meteorology, 84: 153-167. DOI:10.1016/S0168-1923(96)02383-0 |