 2010, Vol. 46
2010, Vol. 46文章信息
- 诸葛飘飘, 罗森林, 王满囷, 张国安
- Zhuge Piaopiao, Luo Senlin, Wang Manqun, Zhang Guoan
- 云斑天牛头部附器感器的扫描电镜观察
- Observation of Sensilla on the Cephalic Appendages of Batocera horsfieldi with Scanning Electron Microscope
- 林业科学, 2010, 46(5): 116-121.
- Scientia Silvae Sinicae, 2010, 46(5): 116-121.
- DOI: 10.11707/j.1001-7488.20100519
-
文章历史
- 收稿日期:2009-04-16
-
作者相关文章



云斑天牛(Batocera horsfieldi),属鞘翅目(Coleoptera),天牛科(Cerambycidae),广泛分布于我国的大片省区,是多种林木的重要蛀干害虫(高瑞桐等,1995;严敖金等,1997)。由于天牛是一类保护机制较为完善的害虫,一般的化学防治很难奏效。目前基于天牛化学生态学及行为研究的防治技术已成为探索天牛防治的新途径。研究发现,天牛感受化合物的主要器官为触角、下颚须和下唇须,上面密布形态与功能各异的感器,这些感器与天牛取食、交配、产卵等活动关系密切(陆群等,2002;韩颖等,2005;江望锦等,2005;程红,2006;诸葛飘飘等,2009)。云斑天牛成虫对寄主植物气味行为反应的研究也表明:嗅觉在云斑天牛寄主定位过程中起重要的作用(李娟等,2008),但对其感受器的研究还未见相关报道。为此,本试验对云斑天牛成虫的触角、下颚须和下唇须进行扫描电镜观察,为进一步探索其行为并利用行为来防治天牛提供依据。
1 材料与方法 1.1 试验昆虫云斑天牛成虫采自湖北省公安县。
1.2 试验方法 1.2.1 基本形态观察触角、下颚须和下唇须的长度及基本特征在解剖镜下进行测量和观察,分别测试15头雌、雄成虫鲜标本的体长和头部各附肢的长度,雌雄之间的差异采用t检验进行比较。
1.2.2 扫描电镜观察将云斑天牛成虫的触角、下颚须和下唇须分别从基部切下,浸于0.1 mol·L-1 PBS缓冲液(pH7.4)中,用KQ-250B型超声波清洗器清洗10 min,再置1%碘化钾组织导电液中浸泡1天,用蒸馏水清洗。自然干燥后,按雌、雄、背面、腹面分别用导电胶将样固定在样品台上,JFC-1600型离子溅射仪喷金,在JSM-6390LV型扫描电子显微镜下观察和拍照。电镜工作电压为20 kV。感受器的类型主要参考Schneider(1964)、Altner(1977)和Zacharuk(1985)对触角感器的命名系统进行命名。
1.2.3 触角感器的形态大小及密度统计由于云斑天牛成虫个体大,触角长,感器数量繁多,不能逐一统计,腹面比背面感器密集,因此采用单位面积法,计算触角腹面每105 μm2面积内各种类型感器的数量以此来反应整体的变化趋势。同时采用t检验对各类感器的长度、分布等在雌雄之间的差异进行分析。
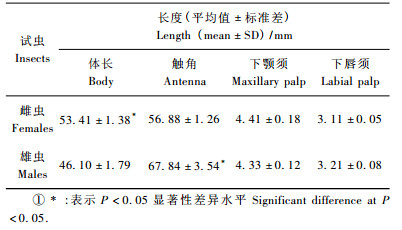
2 结果与分析 2.1 触角、下颚须和下唇须的一般形态云斑天牛两性成虫触角均为线状,着生在复眼间,角质化程度高,由11节组成:柄节(scape)1节,梗节(pedicel)1节,鞭节(flagellomere)9个亚节。柄节粗圆,近似椭圆形;梗节是触角中最为短小的一节,近似球形;鞭节的第1亚节最长,朝末端方向依次变短变细。下颚须4节,下唇须3节,两者外形相似,第1节短小,末节膨大,顶端呈圆锥状,各节段的基部包埋在上一节段端部之内,形成关节状构造,柔软可弯曲。测量结果显示,雌虫体长(53.41±1.38)mm,显著大于雄虫(46.1±1.79)mm,但雄虫触角(67.84 mm±3.54 mm)却显著长于雌虫触角(56.8 mm±81.26 mm)(P<0.05);下颚须长4.2~4.6 mm,下唇须长3.0~3.3 mm,在两性之间不存在显著差异(表 1)。
|
|
电镜下观察,云斑天牛成虫的触角表面具波形刻纹,越靠近触角的末端刻纹越明显。触角感器的数量、类型从柄节到末鞭节逐渐增多。1—6鞭节端部都有一段不着生感器的空白区域,且向端部空白区逐渐变窄,7—9鞭节无空白区。根据感器的形态特征对云斑天牛雌雄成虫触角上的感器进行分类,可分为5个类型:毛形感器、锥形感器(5个亚型)、刺形感器(2个亚型)、凹槽钉形感器、乳突形感器。除此之外,触角表皮上还分布着大量的表皮孔和一些特殊的表皮结构。
2.2.1 毛形感器(sensillum trichodea,T)毛形感器着生于触角表面的圆形凹窝内,感器壁表面具倒“V”字形纵脊,基窝内缘有2个对称的嗅孔。感器从基部着生处向端部逐渐变细,状似毛发,柔软纤长,匍匐于触角表面形成约30°角,有的在近端处向触角的外围空间弯曲,有的从基部弯曲(图版Ⅰ-1)。毛形感器长约77 μm,直径约8 μm,其形态在雌雄间不存在差异(图 1)。毛形感器在整个触角上均有分布,数量从柄节至鞭节的最后一节逐渐增加,且腹面比背面密集,是触角表面分布最广、数量最多的感器。就整体分布密度而言,雌雄成虫间没有差异(图 2)。

|
图版Ⅰ Plate Ⅰ 1.触角的毛形感器T; 2.触角的粗长锥感器BⅠ; 3.触角的粗短锥感器BⅡ、发锥感器BⅢ和指锥形感器BⅣ; 4.触角的指锥形感器B Ⅳ; 5.触角的角锥形感器BⅤ; 6.触角梗节的短刺形感器CHⅠ; 7.触角的长刺形感器CHⅡ; 8.触角梗节的凹槽钉形感器Gp。 1. Sensillum trichodea (T) on the antenna; 2. Sensillum basiconcaⅠ (BⅠ) on the antenna; 3. Sensillum basiconca Ⅱ (BⅡ), sensillum basiconca Ⅲ(BⅢ) and sensillum basiconca Ⅳ(BⅣ) on the antenna; 4. Sensillum basiconca Ⅳ(BⅣ) on the antenna; 5. Sensillum basiconcaⅤ (BⅤ) on the antenna; 6. Sensillum chaeticaⅠ(ChⅠ) on the antenna; 7. Sensillum chaeticaⅡ(ChⅡ) on the antenna; 8. Sensillum grooved peg (Gp) on the antenna. |

|
图 1 云斑天牛雌雄成虫触角上各类感器的长度 Figure 1 The mean length of antennal sensilla in Batocera horsfieldi of each sexs *:表示P<0.05显著性差异水平。 Means significant difference at P < 0.05. |

|
图 2 几种主要感器在云斑天牛雌雄成虫触角各节段腹面的分布数量 Figure 2 Number and distribution of several primary sensilla in abdominal antennal segments of female and male Batocera horsfieldi Sc:柄节Scape; P:梗节Pedice; F1-F9表示鞭节的第1~9亚节First-ninth flagellomere. |
锥形感器广泛分布于鞭节,极少数出现在梗节,柄节上无分布。其基部为圆台形或土丘状隆起的底座,上面着生的锥体形态各异。根据其形态,可以分为5个亚型:
粗长锥形感器(BⅠ)着生于土丘状隆起的底座上,锥体粗大黑色,顶端钝圆,感器壁表面有纵脊,有的端部稍弯,感器长约200 μm,基部直径约35 μm,远粗于其他感器(图版Ⅰ-2),少数分布在梗节背面,分布在各鞭节表面,并有规律地成排环生于这些节段的端部。该感器是唯一在梗节分布的锥形感器,长度在雌雄之间无差异(图 1),但雌虫触角上的数量多于雄虫(图 2)。
粗短锥形感器(BⅡ)形态与粗长锥形感器相似,但锥体明显短许多,且感器壁表面光滑。小锥高约70 μm,基部直径约32 μm(图版Ⅰ-3),长度在两性之间无差别(图 1)。该感器零散分布于鞭节上,数量不多。在雌虫触角上的分布多于雄虫(图 2)。
发锥形感器(BⅢ)着生于圆台形底座,锥体细长,顶端尖,形似发丝。锥高约73 μm,基部直径约8 μm,底座直径约为基部直径的2倍(图版Ⅰ-3)。该感器分布于鞭节的第2~9亚节,与毛形感器相间生长,是触角上数量最多的一种锥形感器,其分布密度仅次于毛形感器。雄虫触角上该感器的数量多于雌虫(图 2),但长度不存在性别之间的差异(图 1)。
指锥形感器(BⅣ)顶端钝圆,直立或微弯,形似翘起的手指,长约25~28 μm(图版Ⅰ-4),也是一种数量丰富的锥形感器。其分布密度在第3-7亚节间无明显差异,但第8~9亚节数量骤然增多。其长度及分布密度在两性之间无差异(图 2)。
角锥形感器(BⅤ)着生于圆台形底座,角状小锥,顶端钝圆,长约7~8 μm,分布于鞭节的第3~9亚节,且数量朝末端方向递增。该感器多集中出现在由鞭节表皮内陷形成的特殊凹窝内,形成锥丛结构(图版Ⅰ-5);其长度在两性之间无差异(图 1),但雄虫上感器数量略多于雌虫(图 2)。
2.2.3 刺形感器(sensillum chaetica,Ch)刺形感器壁表面光滑,顶端尖锐,比毛形感器坚硬,有较强的刚性。根据形态特征,分为2个亚型:
短刺形感器(ChⅠ)直立着生于触角表面,基部明显陷入深窝中(图版Ⅰ-6),雌虫触角上该类感器的长度明显长于雄虫(P<0.05)(图 1)。该感器成簇分布于雌雄成虫梗节侧面和腹面,柄节上也有零星分布,鞭节上无分布,但其分布密度在雌雄间无差异(图 2)。
长刺形感器(ChⅡ)细而长,基窝浅,感器直径约10 μm,长约390~480 μm,明显长于其他感器许多(图版Ⅰ-7),且雌虫触角上该感器的长度显著长于雄虫(图 1),分布密度也显著多于雄虫(图 2)(P<0.05)。该感器在触角各节上均有分布,侧、腹面多于背面,并且有规律地环生于雌雄成虫触角梗节和各鞭节端部,贴近或接触到下一鞭节。
2.2.4 凹槽钉形感器(sensillum grooved peg,Gp)该感器形似钉头,长约35 μm,基部直径约8 μm,从中间弯曲,顶端尖,着生于触角表面的凹窝中,凹窝宽而浅(图版Ⅰ-8)。这种感器只集中分布于梗节腹面的基部。感器的长度和数量在雌雄间无差异(图 1和图 2)。
2.2.5 乳突形感器(sensillum cavitata-peg,Cp)感器突起部分呈乳突状,端部钝圆,长约2.4 μm,基部膨大,直径约14 μm(图版Ⅱ-1)。该感器目前只在雌虫触角鞭节上观察到,且数量少。

|
图版Ⅱ Plate Ⅱ 1.雌虫触角的乳突状感器Cp; 2.下颚须末端的毛形感器T、隙缝感器A和末稍锥形感器TB; 3.下唇须末端末稍锥形感器TB; 4.钟形感器Ca; 5.下颚须末稍锥形感器TBⅠ和TBⅡ; 6.下唇须的末稍锥形感器TBⅡ。 1. Sensillum cavitata-peg(Cp) on female antenna; 2. Sensillum trichodea (T), sensillum aperture (A) and sensillum twig basiconca (TB) on the maxillary palp; 3. Sensillum twig basiconca (TB) on the labial palp; 4. Sensillum campullacea(Ca); 5 Sensillum twig basiconca Ⅰ and Ⅱ (TBⅠ and TBⅡ) on the maxillary palp; 6. Sensillum twig basiconcaⅡ (TBⅡ) on the labial palp. |
下颚须和下唇须上发现的感器类型一样,共5种:毛形感器、长刺形感器、末梢锥形感器、钟形感器和隙缝感器。其毛形感器及长刺形感器的外观形态特征与触角上的相似。毛形感器广泛分布在颚、唇须表面,数量逐节递增,腹面明显多于背面,是数量最丰富的感器。长刺形感器只零星着生在节间衔接处,贴近或接触到下一节,端节上无分布。就整体分布密度而言,这2种感器在雌雄成虫间均无差异,但明显比触角少。
2.3.1 末梢锥形感器(sensillum twig basiconca,TB)这类感器在下颚须和下唇须末端均有分布,形态上无区别。下颚须末节顶端内陷形成一个直径约为120 μm的圆盘(图版Ⅱ-2),其内集中分布约150~180个末梢锥形感器;而下唇须末节顶端钝圆微突(图版Ⅱ-3),没有凹陷,分布约130~150个末梢锥形感器,数量略少于下颚须。
末梢锥形感器短小林立,分布密集,基部着生处有隆起的底座,长约8~12 μm,基部直径约4~5 μm。这类感器从外观上可分为2种:Ⅰ型感器(TBⅠ)顶端中间向内凹陷,外观像火山口(图版Ⅱ-5);主要环生于末梢外缘;Ⅱ型感器(TBⅡ)基部粗,接近顶端处突然缢缩变尖,有多道缢痕(图版Ⅱ-6)。两者间或分布在末梢。形态上,雌雄成虫间无差异;数量上,雌虫多于雄虫。
2.3.2 钟形感器(sensillum campullacea,Ca)钟形感器呈吊钟状突起,着生在大而深的凹窝内,感器体短小,长约2.4 μm,几乎与凹窝边界齐平,基部直径约2 μm(图版Ⅱ-4)。下颚须和下唇须的末梢边缘均有分布,其数量在雌雄成虫间不存在差异,但下颚须多于下唇须。
2.3.3 隙缝感器(sensillum aperture,A)隙缝感器外形好似一道道纵向的缝隙,中间微微凹陷,呈狭长形浅穴,长约28 μm,宽约2.5 μm(图版Ⅱ-2),环绕分布在下颚须和下唇须靠近末端的部位,中间无其他感器。下颚须上的数量比下唇须多,但雌雄成虫间无差异。
3 讨论本研究结果表明:云斑天牛成虫头部附器的外观形态及其感器种类、形态和分布特点与国内外已报道的近缘种天牛存在一定程度的共性,尤其是与其他杨树天牛相似,只是有些感器的命名法不同。云斑天牛成虫触角上共发现了5类感器,其中毛形感器、刺形感器和锥形感器是昆虫触角上的常见感器。毛形感器一般被认为具有感受机械刺激和化学物质的双重功能(马瑞燕等,2000),是目前研究最为广泛的感器之一。在一些鳞翅目昆虫中,毛形感器还具有感受性信息素的作用(Isidoro et al., 1992;Saǐd et al., 2003)。象鼻虫(Isidoro et al., 1992;Saǐd et al., 2003)、小蠹虫(Faucheux,1994)、步甲(Kim et al., 1996)和天牛(Lopes et al., 2002)等鞘翅目昆虫的毛形感器也被陆续证实具备嗅觉功能。感器表面特征往往与其功能相适应,通常可通过微观形态特征来推测功能。云斑天牛的毛形感器壁表面有明显纵脊,着生处有2个嗅孔,这是典型的沟纹型多孔化感器的特征,已知这类感器起嗅觉作用。此外,与多数杨树天牛一致,毛形感器的数量在云斑天牛触角上也占绝对优势,因此可以推测毛形感器是云斑天牛最基础的嗅觉感器,在对寄主气味的嗅觉中起重要作用,但能否识别特异成分还有待进一步深入研究。
基于刺形感器刚性强、长度一般远长于其他感器、能够优先接触物品等特性,多数人认为它的主要功能是机械作用(马瑞燕等,2000;Ochieng et al., 2000)或感受机械刺激和接触性化学刺激(Isidoro et al., 1992)。云斑天牛两性触角上的刺形感器数量没有显著差异,但形态上却有不同,雌虫的刺形感器长度比雄虫长,这说明云斑天牛雌、雄成虫对外界机械刺激的灵敏度可能有所差异,为各自选择适宜的行为环境提供辅助作用。
云斑天牛触角上的锥形感器形态多样,包括5种亚型,与之类似的还有青杨脊虎天牛(Xylotrechus rusticus)(程红,2006)、桑天牛(Apriona germari)(诸葛飘飘等,2009)和桉嗜木天牛(Phoracantha semipunctata)(Lopes et al., 2002)等,这些天牛触角上的锥形感器也都是根据形态差异而划分出多个亚型的。其中,云斑天牛的粗短锥形感器和指锥形感器的外观与青杨脊虎天牛的粗锥形感器和松墨天牛的指形感器(陆群等,2002)相似;角锥形感器聚集形成的锥丛结构也可在桉嗜木天牛(Lopes et al., 2002)和青杨脊虎天牛(程红,2006)等触角上找到类似结构。目前许多学者认为锥形感器起嗅觉作用,有一个宽的、有时重叠的感觉谱,可能是植食性昆虫感受寄主挥发信息素的主要感器(马瑞燕等,2000)。特殊的锥丛模式可以扩大气味感觉区的面积,能够提高触角对气味分子的捕获效率(Lopes et al., 2002),有利于长距离的气味定向(Werner,1995)。云斑天牛锥形感器主要分布于鞭节,从F1到F9的变化趋势是数量逐节增多,亚型也越多,由此可以推测触角的端部是感受化学信息物质的敏感部位,各亚型锥形感器负责的功能上也会有所不同,比如分工识别不同种类的化学物质,而不仅仅只是外部形态上的差异。
云斑天牛成虫下颚须和下唇须共具有5种感器,其形态及感器类型与桑天牛(诸葛飘飘等,2009)、狭胸天牛(Philus antennatus)(尹新明,1994)和黄斑星天牛(Anoplophora nobilis)(李新岗等,1991)等接近,这说明天牛科下颚须和下唇须的基本功能很可能是类似的,不同种之间的形态分化没有太大差异。云斑天牛下颚须和下唇须的顶端末梢分别为一“凹”一“凸”的结构,这有助于增加与气味分子接触的表面积;同时,只集中分布末梢锥形感器这一类感器,也就意味着功能趋向于专一化,可有效提高对某种或某类气味分子的捕获和感知能力。此外,它们的最后一节是感器密集分布区,感器种类和数量都达到最多,这说明下颚须和下唇须的端部是嗅觉和味觉的主要感受区域。早期有研究表明,天牛取食时用末端接触食物可能有“品尝”食物味道的作用,交配时舔翅可能有感受性信息素的作用(尹新明,1994;李新岗等,1991)。
综合比较云斑天牛雌、雄成虫头部附器的外观形态及其超微结构,可知下颚须和下唇须在两性间没有太大差异,但触角存在明显的性二型现象:1)雄虫体型明显小于雌虫,但雄虫触角却显著长于雌虫触角;2)乳突形感器只在雌虫触角上发现,且雌虫触角上的粗长锥形感器(BⅠ)、粗短锥形感器(BⅡ)和长刺形感器(ChⅡ)在数量上多于雄虫,但发锥形感器(BⅢ)少于雄虫;3)雌虫触角上的刺形感器(包括ChⅠ和ChⅡ)的长度显著长于雄虫。针对触角的性别差异,笔者认为应从云斑天牛两性成虫的生活习性的不同点来解释分析。以湖北公安的云斑天牛为例,观察发现成虫从杨树羽化而出,不论雌雄都要到野蔷薇(Rosa multiflora)上补充营养,雌虫交配后又需要回到杨树上产卵;而交配行为都是以雄虫为主动。因此,推测感器类型、分布模式及数量的不同可能会使雌、雄成虫对化学物质的敏感度不同,导致它们对配偶或寄主植物的选择产生细微分化。雌虫触角上特有的乳突形感器以及数量比雄虫多的BⅠ感器、BⅡ感器和ChⅡ感器都可能与产卵寄主的定位选择有关。雄虫拥有更长的触角可能与它依靠触角来接触、定位和识别雌虫的习性相关,有利于提高择偶效率(Hanks,1999)。此外,雄虫触角的锥形感器总量低于雌虫,但发锥形感器BⅢ的数量却明显多于雌虫,暗示发锥形感器可能是用来感受性信息素的关键感器。当然这些结论和尚未明确的问题还需要对感器进一步做电生理研究来验证。
程红. 2006. 青杨脊虎天牛触角感器类型及其对植物挥发物的反应. 哈尔滨: 东北林业大学林学院硕士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-10225-2006111153.htm
|
高瑞桐, 王宏乾, 徐邦新, 等. 1995. 云斑天牛补充营养习性及与寄主树关系的研究[J]. 林业科学研究, 8(6): 619-623. |
韩颖, 张青文, 路大光. 2005. 松褐天牛触角感器的扫描电镜观察[J]. 昆虫知识, 42(6): 681-685. DOI:10.7679/j.issn.2095-1353.2005.170 |
江望锦, 嵇保中, 刘曙雯, 等. 2005. 天牛成虫信息素及嗅觉感受机制研究进展[J]. 昆虫学报, 48(3): 427-436. |
李娟, 王满囷, 张志春, 等. 2008. 云斑天牛成虫对植物气味的行为反应[J]. 林业科学, 44(6): 165-170. |
李新岗, 张克斌. 1991. 黄斑星天牛成虫下颚须和下唇须的化学感受器[J]. 昆虫知识, 28(6): 357-358. |
陆群, 张玉凤. 2002. 光肩星天牛成虫头部附器超微结构的研究分析[J]. 内蒙古林业科技, (1): 16-19. |
马瑞燕, 杜家纬. 2000. 昆虫的触角感器[J]. 昆虫知识, 37(3): 179-183. DOI:10.7679/j.issn.2095-1353.2000.041 |
严敖金, 嵇保中, 钱范俊. 1997. 云斑天牛Batocera horsfieldi (Hope)的研究[J]. 南京林业大学学报, 21(1): 1-6. |
尹新明. 1994. 狭胸天牛成虫下颚须和下唇须化感器研究[J]. 西南农业大学学报, 16(3): 270-271. |
诸葛飘飘, 葛红梅, 王满囷, 等. 2009. 桑天牛头部附器感器的扫描电镜观察[J]. 昆虫知识, 46(2): 238-243. DOI:10.7679/j.issn.2095-1353.2009.042 |
Altner H. 1977. Insect sensillum specificity and structure: an approach to a new typology//Le Magnen J, MacLeod P. Olfaction and Taste Ⅳ, Information Retrieval, London, 295-303.
|
Faucheux M J, Kristensen N P, Yen S H. 2006. The antennae of neopseustid moths: Morphology and phylogenetic implications, with special reference to the sensilla (Insecta, Lepidoptera, Neopseustidae)[J]. Zoologischer Anzeiger, 245(2): 131-s. DOI:10.1016/j.jcz.2006.05.004 |
Faucheux M J. 1994. Distribution and abundance of antennal sensilla from two populations of the pine engraver beetle, Ips pini (Say) (Coleoptera: Scolytidae)[J]. Annales des Sciences Naturelles: Zoologie et Biologie Animale, 15(1): 15-31. |
Hanks L M. 1999. Influence of the larval host plant on reproductive strategies of cerambycid beetles[J]. Annual Review of Entomology, 44: 483-505. DOI:10.1146/annurev.ento.44.1.483 |
Isidoro N, Solinas M. 1992. Functional morphology of the antennal chemosensilla of Ceutorhynchus assimilis Payk (Coleoptera: Curculionidae)[J]. Entomologica, 27: 69-84. |
Kim J L, Yamasaki T. 1996. Sensilla of Carabus (Isiocarabus) fiduciarius saishutoicus Csiki (Coleoptera: Carabidae)[J]. International Journal of Insect Morphology and Embryology, 25(1-2): 153-172. DOI:10.1016/0020-7322(95)00015-1 |
Lopes O, Barata E N, Mustaparta H, et al. 2002. Fine structure of antennal sensilla basiconica and their detection of plant volatiles in the Eucalyptus woodborer, Phoracantha semipunctata Fabricius (Coleoptera: Cerambycidae)[J]. Arthropod Structure and Development, 31(1): 1-13. DOI:10.1016/S1467-8039(02)00011-7 |
Ochieng S A, Park K C, Zhu J W, et al. 2000. Functional morphology of antennae chemoreceptors of the parasitoid Microplitis croceipes (Hymenoptera: Braconidae)[J]. Arthropod Structure and Development, 29(3): 231-240. DOI:10.1016/S1467-8039(01)00008-1 |
Saǐd I, Tauban D, Renou M, et al. 2003. Structure and function of the antennal sensilla of the palm weevil Rhynchophorus palmarum (Coleoptera.Curculionidae)[J]. Journal of Insect Physiology, 49(9): 857-872. DOI:10.1016/S0022-1910(03)00137-9 |
Schneider D. 1964. Insect antennae[J]. Annual Review of Entomology, 9: 103-122. DOI:10.1146/annurev.en.09.010164.000535 |
Werner R A. 1995. Toxicity and repellency of 4-allylanisole and monoterpenes from white spruce and tamarack to the spruce beetle and eastern larch beetle (Coleoptera: Scolytidae)[J]. Environmental Entomology, 24(2): 372-379. DOI:10.1093/ee/24.2.372 |
Zacharuk R Y. 1985.Antennae and sensilla//Kerkut G A, Gilber L I. Comparative Insect Physiology, Biochemistry and Pharmacology. Pergamon Press, Oxford, 6: 1-69.
|