 2010, Vol. 46
2010, Vol. 46文章信息
- 周亮, 刘盛全, 刘亚梅, 刘群燕
- Zhou Liang, Liu Shengquan, Liu Yamei, Liu Qunyan
- 欧美杨107杨偏心生长应变分布规律
- Distribution Pattern of Growth Strain of Eccentric Growth Poplar 107 Clone Tree (Populus ×euramericana cv.'Neva')
- 林业科学, 2010, 46(4): 171-177.
- Scientia Silvae Sinicae, 2010, 46(4): 171-177.
-
文章历史
- 收稿日期:2008-12-11
-
作者相关文章



树木在生长过程中容易受到许多外界因素的干扰,如地形、风吹、雪压、相临木竞争等,使得树木偏离原有的生长状态,发生树干倾斜、弯曲、树冠偏冠等现象(Alméras et al., 2005; John et al., 2003; Yamashita et al., 2007)。在这些因素的干扰下,树木为了更好地获得生存空间必须形成一个抵御机制。立木中生长应力的存在及其分布方式是其抵御外界因素的主要机制之一。生长应力产生于树木次生生长过程,是形成层活动期内分化出来的细胞在发育成熟过程中细胞间产生的相互作用力,可以随着树木的径向生长逐渐积累形成具有一定特定分布方式的应力场。这种树木内部应力会调整在树木中大小和分布方式,进行自我调节来抵御外力并尽力恢复到原有生长状态。当外界干扰因素达到一定程度后,树木还会通过偏心生长形成应力木来主动调节树干的生长方向,以达到恢复原有状态的目的,从树干形态上则表现为倾斜或者弯曲(Bamber, 1987; Boyd, 1950; Chafe, 1979; Wilson et al., 1996)。对于正常生长的树木,轴向表面生长应力通常为拉应力,弦向表面生长应力表现为压应力,径向表面生长应力几乎为零(Alméras et al., 2005; Archer, 1986)。当立木发生偏心生长时,倾斜树干可能在倾斜树干下部(针叶树种)形成应压木,或在倾斜树干上部(阔叶树种)产生应拉木。应压木区域轴向表面生长应力表现为较低拉应力乃至表现为压应力,应拉木区域轴向表面生长应力表现为高拉应力,其生长应力的大小和分布规律与正常状态下的立木之间存在较大的差异(Huang et al., 2001; Okuyama et al., 1990; 1994; Yamamoto et al., 1991)。
人工林木材已经成为我国木材加工行业的主要原料来源(黎云昆, 2003)。由于立地条件较差(如栽植地坡度较大等)、栽培措施不当等诸多因素的存在,在人工林木材中存在一定量的偏心生长林木(Washusen et al., 2003; 金正道, 2001)。发生偏心生长的立木在砍伐和制材过程中,容易产生立木劈裂、板材扭曲或翘曲、制材时夹锯等不利于木材加工利用的负面状况(Yang et al., 2001)。人工林杨树木材是我国木材加工行业主要原料来源之一(叶克林等, 2003),在人工林杨树木材加工利用中,偏心生长的立木对木材加工利用也经常带来负面影响(Constant et al., 2003; 方长华, 2007),因此本文以目前栽植推广面积较广的欧美杨107杨(Populus ×euramericana cv.‘Neva’)为研究材料(张绮纹等, 2003),研究其偏心生长立木生长应力的大小及其在树干外围和内部的分布规律,以期为偏心生长杨树的加工利用提供一定的理论依据。
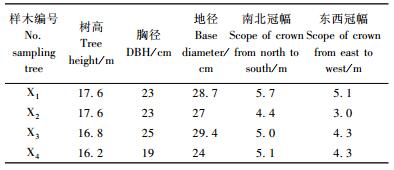
1 材料与方法 1.1 试材采集采集地位于河南省濮阳市白岗乡林场,该林场位于黄河滩区,为黄河冲积平原,土壤类型为轻砂壤,偏盐碱,pH 7.8左右。按照小班数据进行林地踏查,从林地东西向主干渠两旁选择了具有4株7年生偏心生长欧美杨107杨,依次编号为X1, X2, X3, X4,用于测定立木表面轴向生长应变,4株偏心生长欧美杨107杨树干倾斜角度或弯曲程度均不相同,每个单株树形如图 1所示, 样木生长性状如表 1所示。从上述4株平均木中挑选出3株(X1, X3, X4),伐倒后从树干基部开始截成一根4 m长木段,运回实验室进行残余生长应变研究。

|
图 1 4个偏心生长样木树干形态 Figure 1 Shape of trunk of four eccentric growth trees |
|
|
1) 立木表面轴向生长应变测试方法 在立木状态下直接使用应变片法测定倾斜或弯曲树干表面轴向生长应变值。在测试点的选择中,对X1, X2, X3, X4进行了0.5, 1.5, 2.5 m 3个不同高度的测定。在选定的高度上,目测树干倾斜最下部或弯曲最凸部(预测为年轮最窄区域)为定点,将树干外围周长进行7等分,共得到8个等分点,在倾斜最下部或弯曲最凸部为起点,记为0°位置,顺时针对每个等分点进行编号即从0°号点顺时针旋转至315°号点,分别以0°, 45°, 90°, 135°, 180°, 225°, 270°和315°对测定点进行编号,相对应的180°位置为倾斜最上部或弯曲最凹部(预测为年轮最宽窄区域),通过工具在立木表面除去树皮和形成层,得到次生木质部,随后采用应变片法进行生长应变的测定(Yoshida et al., 2000)。
2) 伐倒木内部残余轴向生长应变测试方法 测定材料为3株伐倒木(X1, X3, X4) 4 m长木段的1.15~1.45 m处, 从树干倾斜最下部(0°位置)到最上部(180°位置)或弯曲树干弯曲最凸部(0°位置)和最凹部(180°位置)之间制作径向中心板,保持两端木材完好无损的情况下,在该处制成厚度为4 cm、长度为40 cm、宽度为木段直径的从年轮最宽区域到年轮最窄区域中心板,利用工具剔除其余木材,并将表面砂光,清理干净。用铅笔在中心板上每个年轮的中心位置进行标记,以各年轮中心位置为测试点。从应拉木区(年轮较宽区域)和对应木区(年轮较窄区域)共测试了11个点的内部残余应变,对2个区分别逐年轮编号。对应木区从树皮到髓心逐年轮依次编号为:对7、对6、对5、对4、对3;应拉木区从树皮到髓心逐年轮依次编号为:拉7、拉6、拉5、拉4、拉3。由于第1年轮和第2年轮较窄,所以将其归结一个年轮区域,并在其中心位置,即髓心处标记为1~2。在测试点位置逐年轮平行粘贴应变片,采用应变片法进行残余生长应变测定进行残余轴向生长应变测试(Yoshida et al., 2000)。
3) 数据处理方法 采用SPSS软件对数据进行多因素方差分析。
2 结果与分析 2.1 立木表面轴向生长应变研究本文共测试了4株偏心生长立木的3个不同高度(0.5, 1.5, 2.5 m)周向上8个等分点的立木表面轴向生长应变,其结果如图 2所示, 所测定的立木表面轴向生长应变中,最大值为348 με,最小值为-2 443 με。本文中得出的生长应变的数值大小与其他学者对人工林杨树的研究结果较为一致(方长华,2007),其不同点主要在于X1样木0.5和1.5 m处树干倾斜最下部附近生长应变值为正值,即说明此处产生了拉应变,生长应力表现为压应力。Clair等(2003)用钻孔法测定欧洲毛栗(Casianea sativa)立木表面轴向生长应变时也在倾斜树干的下部发现了压应力。对于这一结果的形成原因将结合残余应变相应研究结果进行详细论述。

|
图 2 X1, X2, X3, X4不同高度立木表面轴向生长应变周向变异模式 Figure 2 Periphery variation curve of surface longitudinal growth strain at different heights of four standing trees X1, X2, X3, X4 |
不同单株、不同高度的立木轴向表面生长应变对不同周向位置作图,其结果如图 2所示。整体上看来,X1, X2, X3, X4立木表面轴向生长应变的周向变异模式在不同单株、不同高度较为一致。从0°位置开始顺时针旋转时,其立木表面轴向生长应变逐渐降低,到达某一位置出现最小值后又开始上升回到原有水平。立木表面轴向生长应变在倾斜树干最上部或弯曲树干最凸处的数值最小,随着位置点在周向上离树干倾斜最下部或弯曲树干最凹处距离的缩小,其应变值逐渐增大。从生长应力角度来看,整个树干外围主要生长应力形式主要以拉应力为主,并且倾斜树干最上部或弯曲树干最凸处的生长应力为拉应力的最大值,倾斜最下部或弯曲树干最凹处的生长应力为拉应力最小值或部分单株出现了压应力,周向上不同位置的生长应力在这两极之间连续性地递增或递减。将测试点位置0~315°依次标号为数字1~8,并以此为自变量对立木表面轴向生长应变进行二次曲线回归,得到4个倾斜树干单株的整体生长应变周向变异模式,结果如图 3所示。由图 3可知,整个应变的周向变异趋势如开口向下的抛物线,顶点在4号和5号位置之间,也即在135~180°位置之间。本文中欧美杨107杨偏心生长木材的立木表面轴向生长应变在周向上的变异模式和整体趋势与学者们对阔叶材偏心生长木材类似研究得出的结论较为一致(Archer, 1986; Okuyama et al., 1990; 1994; Yamamoto et al., 1991)。

|
图 3 偏心生长木材立木表面轴向生长应变周向变异整体趋势 Figure 3 General variation trend of surface longitudinal growth strain of eccentric growth standing tree around trunk periphery |
从图 2显示的结果可以看出,生长应变周向变异模式在各单株、各树干高度的差别主要表现为表面生长应变最低点在周向上的位置,最低点应变数值和应变变化幅度不同。生长应变最低点位置上差异产生的主要原因可能是由于目测的倾斜最下部或弯曲最凸处与实际树干偏心生长年轮最宽处之间并非全部吻合,存在一定的偏差,此外,应拉木存在比例较大区域也可能并非在偏心生长年轮最宽处。因此,生长应变数值最小值在周向上的位置在不同单株和不同高度之间存在一定浮动,尤其在不同单株之间差异较大。在不同高度之间的偏差幅度相对较小,主要在相邻2个位置之间变动,其主要原因可能是偏心生长年轮较宽区域的应拉木分布在高度上具有一定差异性。应变最低点数值和应变变化幅度的差异性可能是由生长性状和树干形态引起。
通过对不同单株和不同高度得出的应变周向变异模式进行综合比较,可以分析出树干形态(倾斜或弯曲的程度)对周向变异模式的影响方式。对同一单株不同高度下的应变值周向变异模式进行比较发现,4个单株任一单株中2.5 m处较其他2个高度处的应变值均较大,应变值变异幅度较小。观察4个单株的树干形态图不难发现(图 1),树干从基本向上树干逐渐开始向一边开始倾斜或者弯曲,当达到一定程度后开始向原有的生长状态逐渐恢复。本文中研究样木在2.5 m处的树干已经开始步入弯曲或倾斜的恢复阶段,其弯曲或倾斜程度低于其他2个树干高度,从而形成了表面应变变异模式比较得出的结果,而0.5和1.5 m之间弯曲或倾斜程度比较结果由于各单株树干形态之间的差异,而在各单株之间不一致,所以这2个高度之间的表面应变变异模式比较也无法得出统一结果。因此认为,同一单株不同树干高度弯曲和倾斜的程度可以影响到其应变的最小值和变异幅度,此外,2.5 m处应变数值较大的原因可能与树冠重力和树干自身重力也存在一定的关系。同一高度不同单株之间周向变异模式比较发现得出的结果之间具有较大差异性,究其原因可能是单株之间的差异较大引起的,尤其是冠幅、胸径、树干等生长因子对生长应变存在一定的影响。但是,2.5 m处在4个单株之间的应变变异幅度差异较大,而且明显看出,随着倾斜或弯曲程度的增加,其变异幅度增加。综合以上观察结果,可以认为弯曲程度或倾斜程度对应变的周向分布存在一定的影响,且随着程度的增加,其应变变化幅度增加,应变数值降低。在上述比较中同样发现,冠幅、树高、胸径、地径等生长性状通过影响树冠重力和树干自重,以及应力积累程度(主要是胸径、地径指标)来影响应变的大小和分布方式。这些因子协同树干形态因子的共同作用,形成了偏心生长立木表面生长应变周向变异较为复杂的局面。因此,需要将正常生长立木表面应变分布研究和偏心生长研究相结合,系统地进行比较和分析,最后才能得出各个影响因子的作用方式。
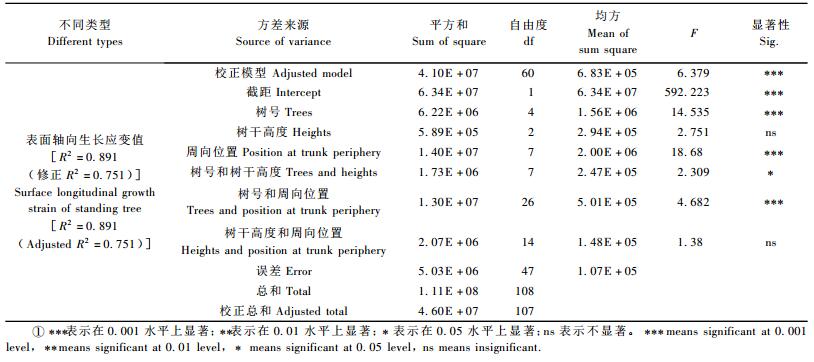
以树号、树干高度、不同周向位置为自变量对表面轴向生长应变值进行三因素方差分析,其结果如表 3所示。三因素方差分析模型在0.001水平上显著,树号和周向位置的主效应,以及树号和周向位置的二元交互效应对生长应变影响程度在0.001水平上显著,树号和树干高度的二元交互效应对生长应变影响在0.05水平上显著,树干高度对生长应变的影响不显著。从方差结果中可知,不同单株和不同周向位置之间的生长应变差异性显著,而同一单株不同高度之间差异不明显。
|
|
将X1, X3, X4伐倒后运回实验室,采用应变片法测试内部残余轴向生长应变径向分布情况, 结果如图 4A所示。3株偏心生长欧美杨107杨伐倒木内部残余轴向生长应变径向变化幅度范围为:-1 763~2 279 με。

|
图 4 偏心生长伐倒木内部残余轴向生长应变径向变化模式(A)和整体趋势(B) Figure 4 Radial variation curve (A) and general trend (B) of inner residual longitudinal growth strain of eccentric growth tree |
各单株内部残余轴向生长应变径向变异模式如图 4A所示。X1从对应木区的最外层一直到髓心位置残余轴向生长应变都为正值,说明生长应力表现为压应力,并且从“对7”到“对3”位置其生长应变变化幅度较少,然后迅速在1-2位置下降;在应拉木区,从1-2位置到“拉5”位置残余轴向生长应变也一直为正值,生长应力表现为压应力,并且持续保持着减小的趋势,在“拉6”和“拉7”位置的残余轴向生长应变才转换成负值,应力表现为拉应力。X1这种残余生长应变的变化模式与前人对阔叶树种偏心生长木材中残余轴向生长应变的轴向变化模式描述存在一定的出入(Okuyama et al., 1990; 1994; Yamamoto et al., 1991)。在前人描述中,应拉木区在树干外围区域表现为生长压应变,即拉应力,随着离髓心距离减少,压应变逐渐转变成拉应变,随后拉应变逐渐增加到最大后开始减小(顶点位置位于应拉木区靠近髓心附近位置),到对应木区后,拉应变仍然保持减少的趋势,直至到临近树干外围转变成压应变,即表现为拉应力,且应拉木区拉应力要大于对应木区。X1与其区别主要在对应木区外围生长应变的变化情况上。前人研究结果表明偏心生长木材的对应木区最外围还有生长拉应力存在,只是数值较少,而本文中的X1却从对应木区的树干最外围(对7位置)开始到髓心处均表现为压应力。在立木表面轴向生长应变的周向变化模式研究中发现,在X1径向“对7”位置相对应的X1树干外围“0°”位置的立木表面周向生长应变也为正值,即生长应力表现为压应力。Clair等(2003)用钻孔法测定欧洲毛栗立木表面轴向生长应变时也在倾斜树干的下部发现了压应力,在其文章还引用其他几位学者在文献中的类似结果,他初步认为树冠对树干的压力可能对这一现象进行一定的解释,然而在随后的研究中Clair等(2006)还发现了在伐倒后的3种阔叶材[杨树(Populus euramericana)、欧洲毛栗、球花森氏藤黄(Symphonia globulifera)]树干表面也出现了生长压应力。结合对其相应位置木材性质的研究,他认为某些偏心生长阔叶材为了更好地恢复原来在生存状态,当无法通过形成应拉木增强在倾斜树干上部或弯曲树干凸处的生长拉应力来独立完成这项工作时,可能会在倾斜树干下部或者弯曲树干凹处形成类似于针叶材应压木的木材构造,从而在树干下部形成生长压应力作为上部拉应力的补偿。
从本文研究结果来看:一方面,X1样木的南北冠幅和东西冠幅均大于其他3株样木,存在由于树冠重力及树干自身重力能部分抵消由树木生长积累起来的表面拉应力的可能性;另一方面,伐倒木中对应木区外围出现生长拉应变的结果也可以说明树干下部可能形成了类似应压木的木材构造,产生生长压应力作为上部生长拉应力的补偿机制。将这2点综合考虑,并与本文研究的其他样木结果进行比较,可以给出以下的推论:偏心生长的阔叶材由于立地条件的不同而形成了不同的生长性状,而不同生长性状的立木所能积累产生的生长应力大小程度也不相同,因此某些偏心生长的阔叶树种可能在特定条件下,当树干上部表面生长拉应力无法满足其恢复原来生存状态需求时,在树干下部表面会形成生长压应力作为补充机制,因此产生了类似样木X1的表面轴向生长应变周向变异模式。此外,这种生长压应力能逐年积累,从而整体影响了偏心生长伐倒木内部残余生长应变径向变异的模式。但是,至于这种生长压应力的产生是否来源与应压木的形成,本文因没有相关研究结果,需要深入研究才能进一步证实。
X3从“拉7”到“拉4”生长压应变逐渐减小,在“拉3”位置处转变成拉应变,随后拉应变一直增加直到“对3”和“对4”位置,拉应变之后开始逐渐减少并在“对5”和“对6”位置之间转变成压应变。X4内部的在“拉7”、“拉6”和“拉5”位置表现为轴向压应变,即拉应力,随后在“拉4”位置表现为拉应变,即压应力,并且达到了拉应变的最大值,然后向对应木区延伸至“对5”位置的过程中拉应变逐渐减小,并最终转变成压应变,“对6”和“对7”位置表现为压应变,但是压应变数值较应拉木区要小的多。X4的径向变异模式与前人相关结果近乎一致(Okuyama et al., 1990; 1994; Yamamoto et al., 1991),而X3的变异模式虽在树干外围区域与前人结果较为相同,但是在树干内部的分布模式存在一定差别,主要是顶点位置不同。X3的顶点位于对应木区,而前人研究结果和本文中X4结果表明顶点位于对应木区靠近髓心的位置。X3的径向变异模式与前人对正常生长阔叶材伐倒木残余轴向生长应变径向分布模式更为相似。这种情况产生的原因可能是由于中心板锯解方向没有横穿年轮最宽区域和年轮最窄区域。从树干外围的轴向应变周向变异模式研究中也发现,并非在树干倾斜最上部或弯曲树干最凸处出现最大的轴向拉应变,这也反映出目测的位置与实际树干内年轮分布的情况存在出入,尤其是X3样木1.5 m处,其拉应变最大值在90°位置,但其径向中心板选择的是横穿0°与180° 2个位置。相比较而言,X4中心板选择的位置基本能横穿年轮最宽区域和年轮最窄区域,因此其径向变异模式与前人研究结果一致。
从径向中心板对应木区最外层年轮一直到应拉木区最外层年轮,以1~11的数字进行编号,然后以这编号对3株倾斜树干的内部残余轴向生长应变进行二次曲线回归,得到其整体径向变异趋势图(图 4B)。3个欧美杨107杨偏心生长木材内部残余轴向生长应变的径向变异整体模式如开口向下的抛物线,顶点在髓心处附近。树干外围主要表现为压应变(拉应力),树干内部表现为拉应变(压应变)。从散点图来看,应拉木区树干外围若干年轮的应变值要小于对应木区相对应的年轮,说明应拉木区树干外围的生长拉应力大于对应木区相对应的位置。
3 结论与讨论所测定立木表面轴向生长应变变化幅度为:-2 443~348 με,其整体周向变异模式为:立木表面轴向生长应变在倾斜树干最上部或弯曲树干最凸处的数值最小(压应变最大,即拉应力最大),随着位置点在周向上离树干倾斜最下部或弯曲树干最凹处距离的缩小,其应变值逐渐增大。多因素方差分析显示,不同单株和不同周向位置之间的生长应变差异性显著,而同一单株不同高度之间差异不明显。
所测定伐倒木内部残余轴向生长应变径向变化幅度范围为:-1 763~2 279 με,其径向变异整体模式如开口向下的抛物线,顶点在髓心处附近,树干外围主要表现为压应变(拉应力),树干内部表现为拉应变(压应力)。从散点图来看,应拉木区树干外围若干年轮的应变值要小于对应木区相对应的年轮,说明应拉木区树干外围的生长拉应力大于对应木区相对的位置。
方长华. 2007. 胶质层对杨树应拉木生长应力和木材性质的影响. 安徽农业大学博士学位论文.
|
金正道. 2001. 我国人工林必须进行集约化经营[J]. 中国林业产业, (10S): 35-37. |
黎云昆. 2003. 我国人工林建设与木材工业的发展[J]. 木材工业, 17(2): 4-7. |
叶克林, 王金林. 2003. 人工林杨树木材的加工利用[J]. 木材工业, 17(1): 5-7, 10. |
张绮纹, 李金花. 2003. 杨树工业用材林新品种[M]. 北京: 中国林业出版社.
|
Alméras T E, Thibaut A, Gril J. 2005. Effect of circumferential heterogeneity of wood maturation strain, modulus of elasticity and radial growth on the regulation of stem orientation in trees[J]. Trees, 19: 457-467. DOI:10.1007/s00468-005-0407-6 |
Archer R R. 1986. Growth stresses and strains in trees[M]. Berlin: Springer Verlag.
|
Bamber R K. 1987. The origin of growth stresses: a rebuttal[J]. IAWA Bulletin, 8: 80-84. DOI:10.1163/22941932-90001032 |
Boyd J D. 1950. Tree growth stress. Ⅲ. The origin of growth stress[J]. Australian Journal of Scientific Research, Series B, Biological Sciences, 3: 294-309. |
Chafe S C. 1979. Growth stress in trees[J]. Australian Forest Research, 9: 203-223. |
Clair B, Ruelle J, Thibaut B. 2003. Relationship between growth stresses, mechanical-physical properties and proportion of fibres with gelatinous layer in chestnut (Castanea sativa Mill)[J]. Holzforschung, 57: 189-195. |
Clair B, Tancrède A, Junji S. 2006. Compression stress in opposite wood of angiosperms: observations in chesnut, mani and poplar[J]. Annals of forest science, 63: 507-510. DOI:10.1051/forest:2006032 |
Constant T, Mothe F, Bradaia M A, et al. 2003. How to relate the standing tree shape to internal wood characteristics: Proposal of an experimental method applied to polar tree[J]. Annals of Forest Science, 60: 371-378. DOI:10.1051/forest:2003028 |
Huang Y S, Chen S S, Lin T P, et al. 2001. Growth stress distribution in leaning trunks of Cryptomeria japonica.[J]. Tree Physiology, 21: 261-266. DOI:10.1093/treephys/21.4.261 |
John R B, George J. 2003. Wood Quality and its Biological Basis. Blackwell Publishing Ltd, Oxford.
|
Okuyama T, Yamamoto H, Iguchi M, et al. 1990. Generation process of growth stresses in cell walls Ⅱ. Growth stresses in tension wood[J]. Mokuzai Gakkaishi, 36: 797-803. |
Okuyama T, Yamamoto H, Yoshida M, et al. 1994. Growth stresses in tension wood : role of microfibrils and lignification[J]. Annals of Forest Science, 51: 291-300. DOI:10.1051/forest:19940308 |
Washusen R, Jugo l, Waugh G. 2003. The relationship between longitudinal growth strain, tree form and tension wood at the stem periphery of ten-to eleven-year-old Eucalyptus globulus Labill[J]. Holzforschung, 67: 308-316. |
Wilson B F, Gartner B L. 1996. Lean in red alder (Alnus rubra): growth stress, tension wood, and righting response[J]. Canadian Journal of Forest Research, 26: 1951-1956. DOI:10.1139/x26-220 |
Yamamoto H, Okuyama T, Yoshida M, et al. 1991. Generation process of growth stresses in cell walls Ⅲ. Growth stresses in compression wood[J]. Mokuzai Gakkaishi,, 37: 94-100. |
Yamashita S, Yoshida M, Takayama S, et al. 2007. Stem-righting mechanism in gymnosperm trees deduced from limitations in compression wood development[J]. Annals of Botany, 99: 487-493. DOI:10.1093/aob/mcl270 |
Yang J L, Waugh G. 2001. Growth stress, its measurement and effects[J]. Australian forestry, 64: 127-135. DOI:10.1080/00049158.2001.10676176 |
Yoshida M, Okuyama T. 2000. Techniques for measuring growth stress on the xylem surface using strain and dial gauges[J]. Holzforschung, 56: 739-746. |