 2010, Vol. 46
2010, Vol. 46文章信息
- 赵燕, 董雯怡, 张志毅, 李吉跃, 聂立水, 沈应柏
- Zhao Yan, Dong Wenyi, Zhang Zhiyi, Li Jiyue, Nie Lishui, Shen Yingbai
- 施肥对毛白杨杂种无性系幼苗生长和光合的影响
- Effects of Fertilization on Seedling Growth and Photosynthesis of Hybrid Clone Seedlings of Populus tomentosa
- 林业科学, 2010, 46(4): 70-77.
- Scientia Silvae Sinicae, 2010, 46(4): 70-77.
-
文章历史
- 收稿日期:2009-07-23
-
作者相关文章



2. 北京林业大学省部共建森林培育与保护教育部重点实验室 100083;
3. 华南农业大学林学院 510642;
4. 北京林业大学水土保持与荒漠化防治教育部重点实验室 100083
2. Key Laboratory for Silviculture and Conservation, Ministry of Education, Beijing Forestry University Beijing 100083;
3. College of Forestry, South China Agriculture University Guangzhou 510642;
4. Key Laboratory of Soil and Water Conservation and Desertification Combating, Ministry of Education, Beijing Forestry University Beijing 100083
毛白杨(Populus tomentosa)是我国北方特有的乡土树种,适应性和抗逆性强,材质优良,在杨树中名列前茅。施肥是培养优质苗木的关键技术环节之一,大量的研究证明,施肥可以促进苗高、地径的生长,增加苗木生物量,提高合格苗产量(刘勇等,2000;Albaugh et al., 2004;Jeyanny et al., 2009),施肥也是培养速生丰产林的关键技术环节之一(孙时轩等,1995),研究表明(陈玉娥等,1998;段树生等,2007),杨树对于土壤的营养条件要求较高,科学施肥能够有效改善土壤条件,增加土壤肥力,促进树木生长。目前关于施肥对毛白杨生长和生理指标的研究有不少报道,刘寿坡等(1988)认为氮肥对毛白杨生长有明显的促进作用,钾肥有一定影响,而磷肥作用不明显;姜岳忠等(2004)研究表明毛白杨苗木对养分的吸收以氮为主,磷、钾次之; 孙时轩等(1995)认为在沙地造林适宜施肥量为450 kg·hm-2,氮磷钾的最佳施肥比例为4:3:0;曹帮华等(2004)通过对毛白杨苗期试验研究得出理想的施肥配方和配比。大量研究表明林木不同基因型的生长对施肥响应的差异较大,应根据不同的基因型配置不同的施肥配方(Li et al., 1991; Jonsson et al., 1997; Miller et al., 2003; McKeand et al., 2003),因此对不同的毛白杨杂种无性系施加相同的肥料是不科学的。本试验主要通过研究不同施肥处理对2个毛白杨杂种无性系苗木生长和光合生理的影响,探索2个无性系的最佳施肥配方,实现生产施肥按需进行、环保经济的目的,从而为针对毛白杨杂种无性系合理施肥提供科学的理论依据。
1 材料与方法 1.1 试验材料供试材料为2个毛白杨杂种无性系:S86和46号。无性系S86和46号都是白杨多交杂种,其母本是毛新杨(Populus tomentasa×P. bolleana),父本是银腺杨(P. alba×P. glandulosa)。2008年3月中下旬将2个毛白杨杂种无性系扦插到37 cm×28 cm(径×高)的塑料桶中,每桶装土16 kg,土壤为沙壤土,基本理化性质:全氮0.303 g·kg-1,速效磷6.62 mg·kg-1,速效钾34.6 mg·kg-1,pH 8.77,密度1.47 g·cm-3,田间持水量14.89%。
氮肥为含N 46.4%的尿素, 磷肥为含P2O5 12.1%的过磷酸钙, 钾肥为含K2O 50%的硫酸钾。
1.2 试验设计采用氮、磷、钾3因素3水平的正交设计(续九如等,1995),按正交表L9(34)进行设计,氮、磷、钾分别放在L9(34)的第1,4,3列上(表 2),对照(CK,不施肥)单列,共10个处理,重复3次,氮磷钾水平及用量(孙时轩等,1995;姜岳忠等,2004)见表 1, 施肥处理见表 2。
|
|

|
|
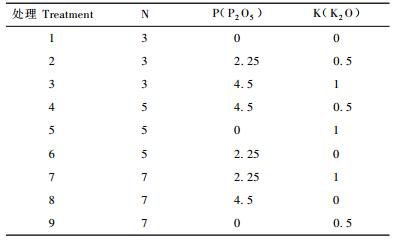
N, P, K在6, 7, 8月中上旬分3次随浇水平均施入土壤中。
1.3 测定方法 1.3.1 苗高、地径及生物量于试验结束时(9月23日)每个处理每个系号选取3株测定苗高、地径及生物量,共测定60株(2个系号,10个处理)。生物量取出后分为根、茎、叶分别称鲜质量,置于烘箱内杀青,70 ℃下烘干至恒重,称干质量。
1.3.2 净光合速率于8月上旬晴朗无云日(8月6日)9: 00—11: 00用Lico-6400测定功能叶片的净光合速率,光强设定为1 000 μmol·m-2s-1,CO2浓度为380~400 μmol·L-1。每个处理测定3个重复。
1.3.3 整株叶面积于8月7日测定整株叶面积,测定株同净光合速率。具体方法为:将叶片分级,每级记录叶片数量,并找出典型叶片用剪纸称重法得出叶面积,以分级叶片数×叶面积再相加之总和得到整株叶面积。
1.3.4 数据处理、分析及图表绘制采用Microsoft Office Excel 2003和SPSS软件(13.0)。
2 结果与分析经过1年的施肥试验,毛白杨无性系S86号和46号的生长情况如表 3所示。
|
|
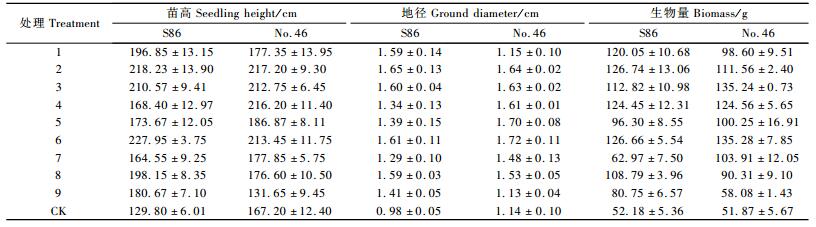
苗高能够反映叶量的多少,体现光合能力和蒸腾面积的大小(沈国舫,2001),因此苗高能够很好反映苗木的生长量。毛白杨杂种无性系S86和46号各处理间苗高方差极显著(表 4),S86各施肥处理均极显著大于对照,其中处理6苗高最大,为227.95 cm(表 3),而对照只有129.80 cm,处理6苗高比对照高75.62%;而处理7苗高最小,为164.55 cm,仅比对照高26.77 %。46号绝大部分施肥处理苗高大于对照,只有处理9苗高低于对照,其苗高为131.65 cm,而对照为167.20 cm,处理9仅为对照的78.74%,可能是由于处理9为不施磷处理,仅是氮、钾配施,苗木营养不均衡从而影响苗高生长所致。
|
|

N, P, K 3因素对苗高影响的平均值与施肥水平建立简单的对应关系见图 1。从图 1中可以看出: S86和46号由氮素引起的苗高变化幅度均为最大, S86由钾素引起的变化幅度次之,减小的速率略小于氮肥;而46号由磷素引起的变化幅度次之,随着磷肥施用量的增加,苗高呈现较大幅度增加后趋于平缓,略微减小的趋势。S86磷肥变化幅度最小,随着磷肥施用量的不断增加,其对苗高的影响由正效应变为负效应,46号钾肥变化幅度最小。由此可见:氮、磷、钾肥对毛白杨无性系S86和46号的苗高影响顺序分别为氮>钾>磷和氮>磷>钾。

|
图 1 不同氮磷钾水平对毛白杨无性系S86号和46号苗高的影响 Figure 1 Effects of N, P, K at different levels on seedling height in Populus tomentosa clone: S86, No.46 |
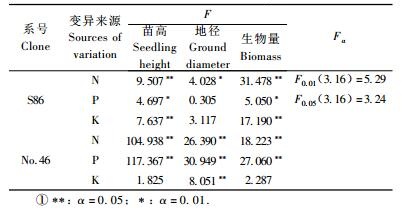
对施肥处理进行正交方差分析,结果见表 5。氮、钾对S86苗高生长影响极显著,这说明氮、钾肥能够很好地促进S86苗高的生长;磷对其苗高生长影响显著,说明磷的作用也很重要。氮、磷对46号苗高生长影响极显著, 这说明氮、磷肥能够很好地促进46号苗高的生长,钾对苗高的生长无显著影响。为了选出最佳水平,对于苗高生长影响显著的元素进行多重比较(表 6),S86号氮水平1(3 g)和2(5 g)差异显著,水平1和3(7 g)差异极显著, 可认为S86氮的最佳施肥水平为1(3 g);而46号氮水平1和2无显著差异,水平1和2与2和3差异极显著,可认为氮的最佳水平为1(3 g)和2(5 g)。S86磷水平1(0 g)和2(2.25 g)差异极显著,水平1和3(4.5 g)与2和3均无显著差异, 可认为S86磷的最佳施肥水平为2(2.25 g)和3(4.5 g);46号磷水平1和2与1和3差异极显著,2和3差异不显著,可认为磷的最佳水平是2(2.25 g)和3(4.5 g)。S86钾水平1(0 g)和2(0.5 g)差异显著,水平1和3(1 g)差异极显著, 可认为S86钾的最佳施肥水平也为1(0 g)。而钾的1水平施肥量为0,这说明S86土壤中本身的钾已经能够满足植物生长的需求,施入过量的钾反而会抑制其生长。
|
|
|
|

以上分析表明:氮、磷、钾对S86苗高影响的最佳水平依次为水平1(3 g), 2(2.25 g)和3(4.5 g), 1(0 g);氮、磷对46号苗高影响的最佳水平依次为1(3 g)和2(5 g), 2(2.25 g)和3(4.5 g)。
2.2 地径地径是反映苗木质量最好的指标之一。毛白杨杂种无性系S86和46号经方差分析各处理间地径方差均达到极显著水平(表 4),S86施肥处理4和7显著大于对照,其他施肥处理极显著大于对照,其中施肥处理2地径最大,为1.65 cm,对照为0.98 cm,处理2比对照高68.50%,施肥处理7地径最小,为1.34 cm,仅比对照高31.12%;而46号绝大部分施肥处理地径大于对照,只有处理9低于对照,其地径为1.13 cm,是对照的98.68%,而处理9的苗高亦低于对照,这进一步说明营养均衡搭配有助于毛白杨杂种无性系46号的生长,反之会抑制。
S86和46号的极差值分别为为RN=0.19 cm>RK=0.17 cm>RP=0.05 cm和RN=0.30 cm>RP=0.29 cm>RK=0.14 cm,说明氮、磷、钾对S86地径影响的大小依次分别为氮>钾>磷和氮>磷>钾。而韩恩贤等(2006)对小叶杨(P. simonii)苗期施肥试验表明磷肥在其地径生长过程中起着主导作用,结论不一致可能是由于土壤养分差异或者树种本身的需肥特性不同造成的。
从表 5可以看出:氮对S86地径生长影响显著,磷、钾影响不显著, 说明氮肥更有利于毛白杨无性系S86地径的生长;氮、磷、钾对46号地径生长影响均达到极显著水平。对于地径生长影响显著的元素进行多重比较,结果见表 7,S86氮水平1(3 g)和2(5 g)与1和3(7 g)差异显著,水平2和3差异不显著, 可认为氮的最佳施肥水平为1(3 g);46号氮水平1和2与2和3差异极显著,水平1和3差异显著,可认为氮的最佳水平为2(5 g)。46号磷水平1(0 g)和2(2.25 g)与水平1和3(4.5 g)差异极显著,水平2和3无显著差异,因此磷肥对于46号地径生长影响最优的是水平2(2.25 g)和3(4.5 g);46号钾水平1(0 g)和3(1 g)与2(0.5 g)和3差异极显著,1和2差异不显著,说明钾肥对于46号地径生长影响最优的是水平3(1 g)。因此氮对S86地径影响的最佳水平为1(3 g);氮、磷、钾对46号地径影响的最佳水平依次为2(5 g), 2(2.25 g)和3(4.5 g), 3(1 g)。
|
|

生物量是植物生长发育产生的物质总量,它是反映苗木生产力水平的重要指标之一。毛白杨杂种无性系S86和46号各处理间生物量方差极显著(表 4),S86施肥处理均极显著大于对照,其中处理2和6的生物量最大,分别为126.74和126.66 g,是对照的2.43倍,处理7生物量最小,为62.97 g,仅为对照的1.21倍。46号绝大部分施肥处理生物量极显著大于对照,只有处理9生物量与对照无显著差异,为58.08 g,仅为对照的1.12倍。
S86和46号施氮肥极差均最大,分别为35.70和35.93,其中S86氮的第1水平平均值最大,为119.87 g,其次是钾肥,施P肥影响最小。46号氮的第2水平平均值最大,为120.03 g,其次是磷肥,施钾肥影响最小。
从正交方差分析表 5易看出:氮、钾对S86生物量影响极显著,磷肥影响显著;氮、磷对46号生物量生长影响极显著, 钾对其生物量的生长无显著影响。对影响显著元素进行多重比较,结果见表 8,S86氮水平1(3 g)和3(7 g)与水平2(5 g)和3差异极显著,水平1和2差异不显著, 因此S86对于生物量的影响氮的最佳施肥水平为1(3 g)和2(5 g);46号氮水平1和2无显著差异,水平1和3与2和3差异极显著,可认为46号对于生物量的影响氮的最佳水平为1(3 g)和2(5 g)。S86磷水平1(0 g)和3(4.5 g)差异极显著,水平1和2(2.25 g)与水平2和3无显著差异,可认为S86对于生物量的影响最佳磷水平为2(2.25 g)和3(4.5 g);46号磷水平1和2与1和3差异极显著,2和3差异不显著,可认为46号对于生物量的影响磷的最佳水平是2(2.25 g)和3(4.5 g)。S86钾水平1(0 g)和3(1 g)与2(每根0.5 g)和3差异极显著,水平1和2无显著差异,因此S86对于生物量的影响钾的最好水平为1(0 g)和2(0.5 g)。由此可见,氮、磷、钾对毛白杨杂种无性系S86生物量的影响的大小依次为氮>钾>磷,氮、磷、钾对46号生物量的影响大小依次为氮>磷>钾。氮、钾对S86生物量影响的最佳水平依次为水平1(3 g)和2(5 g)、水平1(0 g)和2(0.5 g);氮、磷对46号生物量影响的最佳水平依次为水平1(3 g)和2(5 g)、水平2(2.25 g)和3(4.5 g)。
|
|

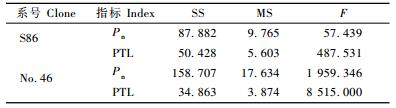
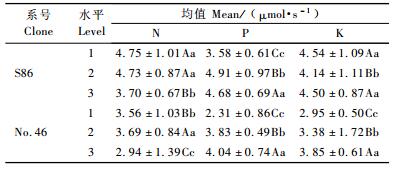
毛白杨杂种无性系S86和46号各处理净光合速率(Pn)和净光合速率×总叶面积(PTL)方差分析均达到极显著水平,其中净光合速率×总叶面积在处理间差异较显著(表 9)。为了进一步探讨这些光合指标与生长的相关关系,利用上述2个光合指标分别与苗高、地径和生物量做一元线性回归,结果见表 10。各个无性系净光合速率×总叶面积与苗高、地径和生物量回归曲线的相关系数均达到了极显著水平,相关系数都在0.800以上,而净光合速率与苗高、地径和生物量回归曲线的相关系数大部分未达到显著水平,只有S86净光合速率与生物量的回归曲线达到显著水平。因此对2个无性系净光合速率×总叶面积进行分析。S86和46号年施肥处理正交方差分析易得到氮、磷、钾对净光合速率×总叶面积影响均达到极显著水平(P<0.01),对其进一步进行多重比较(表 11),S86氮水平1和3与水平2和3差异极显著,水平1和2无显著差异,可认为S86氮的最佳水平为1(3 g)和2(5 g);46号氮各水平之间差异均为极显著,可认为46号氮的最佳水平为2(5 g)。S86和46号磷各水平之间差异均为极显著,可认为S86磷的最佳水平为2(2.25 g),而46号磷的最佳水平为3(4.5 g)。S86钾水平1和3无显著差异,水平1和2与2和3差异极显著,可认为S86钾的最佳水平为1(0 g)和3(1 g);46号钾各水平之间差异极显著,可认为46号钾的最佳水平为3(1 g)。
|
|
|
|

|
|
1) 研究表明:由于毛白杨无性系间本身受较强的遗传因素影响,生长及其生理性状存在较大差异(许兴华等,2006;李爱民等,2005;符军等,1995;王琦等,1995;李继东等,2006;朱春全等,1995),无性系对于肥料的吸收也可能存在差异,通过对毛白杨杂种无性系S86和46号苗高、地径、生物量以及净光合速率×总叶面积指标的综合分析可知: S86最佳施肥处理为N1P2K1,即N:P:K=3:2.25:0,46号最佳施肥处理为N2P3K3, 即N:P:K=5:4.5:1。易发现S86最佳施肥处理对应的总施肥量为5.25 g,46号最佳施肥处理对应的总施肥量为10.5 g,而S86苗高最高可达227.95 cm,46号苗高最高为217.20 cm,S86施肥量较少并且生长较好,因此可以作为节肥优选无性系大量种植推广,从而实现良好的生态效益和经济效益。
2) 林木不同基因型的生长对施肥响应的差异较大,并且其存在交互效应,所以要求根据不同基因型对施肥的反应特性差异进行科学施肥, 实现基因型与施肥的优化配置(Li et al., 1991; Jonsson et al., 1997; McKeand et al., 2003)。McKeand等(2003)的试验表明:不同的火炬松(Pinus taeda)家系对于施肥反应差异很大; 李天芳等(2009)对白桦(Betula platyphylla)7个家系1年生苗木施肥试验结果表明:不同的家系由于其遗传基因不同,其所需要的营养条件不尽相同。本研究对毛白杨杂种无性系S86和46号氮磷钾施肥试验生长及光合指标分析结果表明:不同系号毛白杨苗木对相同施肥处理响应不尽一致,如表 1所示,毛白杨杂种无性系46号对照苗高为167.20 cm,处理9(N:P:K=7:0:0.5)苗高仅为131.65 cm,比对照低21.26 %,而S86对照苗高为129.80 cm,处理9苗高为180.67比其对照高39.18 %。46号可能是由于施肥比例不协调导致处理9比对照苗高低,施肥反而抑制了苗高的生长,而S86该处理比对照高,可见不同的无性系对同一施肥处理响应不同,因此要针对无性系制定不同的施肥配方。
3) 毛白杨无性系各施肥处理苗高、地径、生物量和光合差异均达到极显著水平(表 4, 9)。不同的施肥处理氮、磷、钾的比例不同,氮对光合器官的建成具有重要作用,磷对植株的分蘖、分枝以及根系生长有良好作用,钾是调节植物细胞渗透势的最重要组分,影响植物的蒸腾作用(武维华,2003)。3者在植物体内的作用并非孤立,而是通过有机物的形成与转化得到相互联系。孙时轩等(1995)认为合理的氮、磷、钾比例,能够促使毛白杨林木均衡地吸收营养元素,使叶子合成叶绿素多、光合潜力大,促进毛白杨生长;如果比例失调,根系吸收营养元素则会发生障碍,造成生理活性下降,不利于林木生长。因此在施肥过程中要注意协调氮磷钾施肥比例。
4) 氮元素是植物生长发育所必需的有机氮化合物的构成成分,与植物细胞分裂和生长及整体的生长发育关系密切,大量的研究表明氮元素在施肥过程中对植物的生长影响最大,而磷钾的作用说法不一(曹帮华等,2004;侯传本,2008;姜岳忠等,2004;王洪军,2008;王渭玲等,2008;吴家胜等,2003;武占会等,2004;杨平等,2007;张晓鹏等,2007;周晓舟等,2008)。从本研究结果可知:氮、磷、钾肥对46号和S86生长影响的顺序分别为氮>磷>钾和氮>钾>磷, 其中S86钾的最佳水平为1,即不施钾肥,这说明土壤中的钾已能满足该无性系的生长需要,再施入钾肥反而会抑制其生长。
5) 有研究表明净光合速率与杨树的生长呈正相关(Ceulemans, et al., 1983;1987);而也有研究表明净光合速率与杨树的生长无明显关系,甚至可能出现负相关系数(符军等,1995)。本研究中净光合速率与生长无明显关系, 而净光合速率×总叶面积(PTL)与生长密切相关, 相关系数都在0.80以上, 这可能是由于净光合速率虽然反映了植物积累干物质的能力(李生英等,1998),但是影响植物干物质积累的因子除净光合速率外,还有植物的叶面积,光合产物的运输速度和分配形式等,而净光合速率×总叶面积(PTL)考虑叶面积的差异,更接近于苗木的光合潜力(李静怡等,2000),与生长的相关高于净光合速率与生长的相关,因此,结合净光合速率和叶面积对毛白杨杂种无性系各施肥处理进行评价是比较可靠和理想的。
曹帮华, 巩其亮, 齐清. 2004. 毛白杨杂种苗期不同配方施肥效应的研究[J]. 山东农业大学学报:自然科学版, 35(4): 512-516. |
陈玉娥, 安国英, 牛三义, 等. 1998. 土壤本底与毛白杨幼林施肥效应间关系的研究[J]. 河北林业科技, (2): 1-3. |
段树生, 施海, 王连军. 2007. 施肥对毛白杨生长量、叶片氮(N)、磷(P)、钾(K)含量及蒸腾作用、气孔阻力影响的研究[J]. 广东林业科技, 23(3): 13-17. |
符军, 符毓秦, 刘玉媛, 等. 1995. 田间条件下毛白杨无性系光合和蒸腾特性研究[J]. 陕西林业科技, (6): 1-6. |
韩恩贤, 韩刚, 淮宗安, 等. 2006. 渭北旱原小叶杨苗期的施肥效应[J]. 中南林学院学报, 26(4): 31-36. |
侯传本. 2008. 鲁西潮土小麦玉米氮磷钾配施效应初探[J]. 山东农业科学, (5): 78-79. |
姜岳忠, 吴晓星, 马玲. 2004. 毛白杨苗期生长特性及需肥量研究[J]. 甘肃农业大学学报, 39(4): 423-426. |
李爱民, 赵玉萍, 张英, 等. 2005. 2个毛白杨无性系生长差异性分析[J]. 甘肃林业科技, 30(2): 25-28. |
李继东, 王艳玲, 夏国宗. 2006. 毛白杨无性系叶性状差异及遗传分析[J]. 河南林业科技, 26(3): 12-13. |
李静怡, 张志毅. 2000. 毛白杨杂种无性系光合特性的研究[J]. 北京林业大学学报, 22(6): 12-15. |
李生英, 兰荣光, 许童羽, 等. 1998. 不同杨树品种生理生态特性及其与生长的关系[J]. 沈阳农业大学学报, 29(1): 47-52. |
李天芳, 姜静, 王雷, 等. 2009. 配方施肥对白桦不同家系苗期生长的影响[J]. 林业科学, 45(2): 60-64. DOI:10.11707/j.1001-7488.20090210 |
刘寿坡, 朱占学, 张瑛. 1988. 毛白杨人工林施肥效应的初步研究[J]. 土壤通报, (5): 219-222. |
刘勇, 陈燕, 张志毅, 等. 2000. 不同施肥处理对毛白杨杂种苗木生长及抗寒性的影响[J]. 北京林业大学学报, 22(1): 38-44. |
沈国舫. 2001. 森林培育学[M]. 北京: 中国林业出版社.
|
孙时轩, 张振江, 孙晓雨, 等. 1995. 毛白杨在沙地造林的施肥量及配比(Ⅱ)[J]. 北京林业大学学报, 17(1): 31-36. |
王洪军. 2008. 长白落叶松苗不同配比施肥试验研究[J]. 黑龙江生态工程职业学院学报, 21(5): 38-40. |
王琦, 朱之悌. 1995. 毛白杨优树无性系相关选择研究[J]. 林业科学研究, 8(1): 107-111. |
王渭玲, 王振, 徐福利. 2008. 氮、磷、钾对膜荚黄芪生长发育及有效成分的影响[J]. 中国中药杂志, 33(15): 1802-1806. |
吴家胜, 张往祥, 曹福亮. 2003. 氮磷钾对银杏苗生长和生理特性的影响[J]. 南京林业大学学报:自然科学版, 27(1): 63-66. |
武维华. 2003. 植物生理学[M]. 北京: 科学出版社.
|
武占会, 申书兴, 王惠英, 等. 2004. 天鹰椒氮磷钾高产施肥模型研究[J]. 河北农业大学学报, 27(6): 26-29. |
许兴华, 李霞, 孟宪伟, 等. 2006. 毛白杨优良无性系选育研究[J]. 山东林业科技, (2): 30-32. |
续九如, 黄智慧. 1995. 林业试验设计[M]. 北京: 中国林业出版社.
|
杨平, 孙向阳, 王海燕, 等. 2007. 施肥对台湾青枣营养生长的影响[J]. 北京林业大学学报, 29(6): 211-214. |
张晓鹏, 赵忠, 张博勇. 2007. 氮磷钾对俄罗斯大果沙棘扦插苗生长效应的影响[J]. 西北林学院学报, 22(3): 96-99. |
周晓舟, 唐创业. 2008. 氮磷钾对秋玉米农艺性状和植株养分的影响[J]. 河南农业科学, (9): 27-33. |
朱春全, 王世绩, 王富国, 等. 1995. 六个杨树无性系苗木生长、生物量和光合作用的研究[J]. 林业科学研究, 8(4): 388-394. |
Albaugh T J, Allen H L, Dougherty P M, et al. 2004. Long term growth responses of loblolly pine to optimal nutrient and water resource availability[J]. For Ecol Manage, 192: 3-19. DOI:10.1016/j.foreco.2004.01.002 |
Ceulemans R, Impens I. 1983. Net CO2 exchange rate and shoot growth of young poplar (Populus) clones[J]. J Exp Bot, 34: 866-870. DOI:10.1093/jxb/34.7.866 |
Ceulemans R, Impens U, Steenackers V. 1987. Variations in photosyn-thetic, anatomical, and enzymatic leaf traits and correlations with growth in recently selected Populus hybrids[J]. Can J Bot, 17: 273-283. |
Jeyanny V, Ab Rasip A G, Wan Rasidah K, et al. 2009. Effects of macronutrient deficiencies on the growth and vigour of Khaya ivorensis seedlings[J]. Journal of Tropical Forest Science, 21(2): 73-80. |
Jonsson A, Ericsson T, Eriksson G, et al. 1997. Interfamily variation in nitrogen productivity of Pinus sylvestris seedlings[J]. Scandinavian Journal of Forest Research, 12(1): 1-10. DOI:10.1080/02827589709355377 |
Li B, Mckeand S E, Allen H L. 1991. Genetic variation in nitrogen use efficiency of loblolly pine seedlings[J]. Forest Science, 37(2): 613-626. |
McKeand S E, Grissom J E, Rubilar R, et al. 2003. Responsiveness of diverse families of loblolly pine to fertilization:eight-year results from SETRES-2//Proc 27th South For Tree Impr Conf Stillwater, 30-33. https://www.researchgate.net/publication/241133361_Responsiveness_of_Diverse_Families_of_Loblolly_Pine_to_Fertilization_Eight-Year_Results_from_SETRES-2
|
Miller B D, HawkinsB J. 2003. Nitrogen uptake and utilization by slow-and fast-growing families of interior spruce under constrasting fertility regimes[J]. Canadian Journal of Forest Research, 33(6): 959-966. DOI:10.1139/x03-017 |