 2010, Vol. 46
2010, Vol. 46文章信息
- 任华东, 姚小华, 康文玲, 李生, 王开良, 段福文
- Ren Huadong, Yao Xiaohua, Kang Wenling, Li Sheng, Wang Kailiang, Duan Fuwen
- 黑荆树种源和家系的遗传变异与早期选择
- Genetic Variation and Early Selection of Provenances and Families of Acacia mearnsii
- 林业科学, 2010, 46(3): 153-160.
- Scientia Silvae Sinicae, 2010, 46(3): 153-160.
-
文章历史
- 收稿日期:2008-12-01
-
作者相关文章


2. 云南楚雄林业科学研究所 楚雄 675005
2. Forest Institute of Chuxiong Prefecture, Yunnan Province Chuxiong 675000
黑荆树(Acacia mearnsii)是原产澳大利亚的优良速生多用途树种。20世纪50年代作为优良的栲胶原料引入我国(贺近格等,1991),在我国南方广泛栽培,成为重要的栲胶原料资源,近年来,该树种以其速生性及土壤改良作用,成为适生地区重要的荒山绿化和水土保持树种。种源及家系选择是树木遗传改良的重要手段之一(祝列克, 2001),我国科技人员已对许多树种进行了种源及家系的选择研究(秦国锋等,1997;周志春等,1999;孙晓梅等,2004;郭文福等,2008),结果证明种源家系选择能取得良好的改良效果。我国黑荆树的遗传改良工作始于20世纪80年代初,但系统的研究始于80年代中后期开始实施的中澳合作项目,先后进行了选优、种源选择、家系子代测定等系列研究,但因时间关系,这些研究都只探讨了种源和家系早期生长的遗传变异规律。本文利用15年生的黑荆树自由授粉家系子代测定林幼、中、成3个主要生长阶段的观测资料, 对黑荆树自由授粉家系的树高、胸径、树皮厚度、单株材积等生长性状进行遗传变异分析、遗传参数估计,并对黑荆树自由授粉家系生长性状的早晚相关性及早期选择的效果进行分析,探讨黑荆树家系早期选择的可行性和可靠性。
1 材料与方法 1.1 试验地概况试验地在云南省禄丰县和平乡(102°12′E,25°12′30″N),海拨1 800 m,西南向缓坡,平均坡度10°。属亚热带季风气候,冬夏季短, 春秋季长;日温差大, 年温差小;冬无严寒,夏无酷暑;干湿分明, 雨热同季;日照充足,霜期较短。年平均气温为15~16 ℃,最冷月(1月)平均气温在7.4 ℃,最热月(6月)平均气温21.4 ℃,极端最低温为-5 ℃,年降雨量930 mm,≥10 ℃活动积温5 000 ℃。试验地原为荒坡地,水土流失严重,土壤为白云岩发育的山地红壤,心土裸露,基本上已无熟化的表土,土层厚度大于1.0 m,表层土壤pH 5~6。
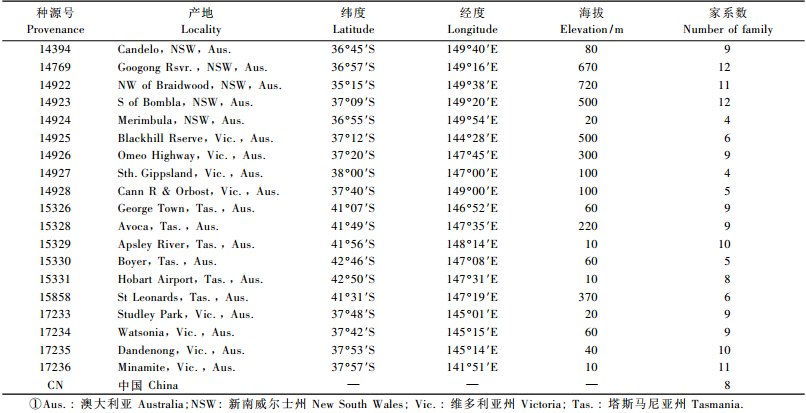
1.2 试验材料与田间设计子代测定林建于1990年7月,采用当年春季培育的容器苗造林。供试材料源自澳大利亚新南威尔士州、维多利亚州和塔斯马尼亚州19个种源区175个半同胞家系及国内早期人工林中选择的8个家系,共计183个家系。
田间试验采用BIB设计,单株小区,20次重复。初植密度2 500株·hm-2,穴状整地(60 cm×60 cm×50 cm)。试验林抚育及管理按常规方式进行。试验林3年生保存率85%以上。至2005年调查时,试验林一直保持自然生长状态,除林木自然竞争导致死亡外,未受任何人为和自然灾害的破坏,总体保存率75%以上。由于试验采用单株小区设计,造林后部分家系保存率较低,小区数据缺失,为保证试验精度和结果可靠性,剔除了15年生时保存率80%以下的17个家系(全部源自澳大利亚种源区),对保存率80%以上的166个家系(表 1)的数据进行统计分析。
|
|
试验林营建后至1995年,每年年终对树高、胸径、冠幅等生长性状进行每木测量,此后分别于1999年和2005年各进行了1次全林调查,其中1993, 1995, 1999和2005年除测量树高、胸径和冠幅外,还进行了树皮厚度的测定。树皮厚度采用2 cm口径的皮带冲在胸高(1.3 m)位置南北两方向取皮样,用游标卡尺测量样皮厚度, 取平均值。单株材积采用二元材积式(林杰等,1987)计算: V=0.000 035 340 508 16×(D+0.7)2.148 743 58×H0.875 969 22,式中: V, D, H分别表示单株材积、胸径和树高。
缺失数据的处理采用Healy等(1956)提出的方法:对缺1个数据的小区按公式Ŷij=(IYi·+JY·j-Y··)/[(I-1)(J-1)]补缺,式中, I为处理数,J为重复数,Ŷij为缺失数据计算值,Yi.为缺失处理所在重复的观测值总和, Y·j为缺失处理所有重复观测值总和; 缺失2个以上的数,除留1个数据外,其他数据先按(yi·+yj·)/2(式中, yi·为缺失处理所在重复观测值均值,yj·为该处理所有重复均值)计算出近似值补缺,然后按单缺株方法进行逐个缺失数据的校正,直到与总体均值残差为零。由于缺失数据对整个方差没有作用,因而在方差分析时总的和误差自由度减去缺失数计算方差。数据补缺由Genstat 5软件Anova分析模块自动计算(Payne et al., 1989)。
1.4 统计分析方法1) 方差及遗传变异分析 种源/家系二水平方差分析采用种源+不等家系分析模型(Snedecor et al., 1980): Yijk=μ+ Pi+Fj+Bk+eijk。式中,Yijk为第i种源第j家系在k重复中的观测值,μ为总体平均值,Pi为种源效应,Fj为种源内家系效应,Bk为重复效应,eijk为机误。
性状的单株遗传力和家系遗传力的计算公式:hi2 = 4σf2/(σe2+σf2); hf2 =σf2/(σe2/r+σf2)。式中, hi2表示单株遗传力;hf2表示家系遗传力;σf2, σe2分别表示家系、环境方差分量,r为试验重复次数。
性状的遗传变异系数和表型变异系数采用下列公式计算: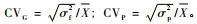
2) 性状早晚相关分析 生长性状的早晚遗传相关、表型相关、环境相关系数和遗传力,均在单株小区值的基础上估算,计算公式: rg(t, T)=σg(t, T)/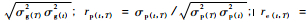
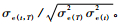
早晚相关遗传增益(ΔG)及早-晚相关遗传进度(RGP)采用如下公式估算: ΔG=ihthTrgtTσpT; RGP=rgtT(ht/hT)。式中: i为选择强度,ht和hT表示早期t和晚期T时的遗传力,rgtT表示时间t-T时的早-晚遗传相关系数,σpT为成熟龄时的表型标准差。
本文试验调查数据采用MS Excel数据处理程序进行整理,方差分析采用Genstat5 (7thed)统计软件的随机区组方差分析模块(Payne et al., 1989)。
2 结果与分析 2.1 家系、种源及种源内家系生长性状表型变异分析对166个黑荆树家系不同林龄主要生长性状变异分析表明(图 1):树高、胸径和树皮厚度家系间差异明显。家系间树高、胸径和树皮厚度的变异系数为14%~22%,不同生长阶段变异系数有所差异,其中胸径与树皮变异系数较大,为19%~20%,而树高变异较小,为14%~16%。对家系间的极差分析也充分说明家系间存在明显的差异,树高、胸径和树皮厚度等生长性状家系间的极差随树龄增加而增加,在生长各阶段最优家系树高生长量及树皮厚度是最差家系的2倍以上,而胸径差异更为显著,最优家系平均胸径是最差家系的3倍以上。15年生时,树高、胸径和树皮厚度的变异幅度分别为6.12~16.07 m, 4.93~17.41 cm和4.18~10.15 mm。为清晰地反映家系间各生长性状的变化趋势,分别对3年生时树高、胸径、树皮厚度均值为高、中、低的各2个家系作生长趋势图(图 2)。从图中可以看出,早期处于生长劣势的家系在整个生长过程中基本上保持劣势状态,不会表现出后期强劲增长的态势,同样,早期生长快的家系也很少表现出后期衰退的趋势。但生长早期处于中等表现的部分家系后期会有突出的表现,特别是胸径生长性状上表现尤为突出,如图 2b中的88号家系,其胸径均值在3年生时仅位居中等,而在5年生时其均值已超过3年生时表现最好的家系,并继续保持优势,这一结果说明以3年生时的材料评价家系优劣为时尚早。

|
图 1 生长性状变异系数的变化趋势 Figure 1 Time trends in variation coefficients (CV) for growth traits BTBH: Bark thickness at breast height.下同。The same below. |

|
图 2 典型家系树高、胸径、皮厚表型均值变化规律 Figure 2 Trends in phenotypic means of height, DBH and BTBH for typical families |
参试家系分属不同的种源,为了解家系间差异的来源及其与种源的关系,对树高、胸径、树皮厚度、单株材积等表型生长性状进行种源和种源内家系间的变异分析(表 2)。结果显示,黑荆树各生长性状种源间及种源内家系间均存在明显的差异,各个生长性状表现最好的种源均值是最差种源均值的1.5倍以上,其中代表综合生长表现的单株材积种源间差异尤为明显,15年生时,最优种源均值是最差种源均值的5倍以上。各生长阶段种源间树高变异系数为0.10~0.12,胸径变异系数为0.15~0.16,树皮厚度早、中期稳定在0.15,但在15年生时,种源间差异有所缩小,变异系数降至0.12,而代表综合生长表现的单株材积种源间的变异系数均在0.30以上;从表 2中的种源内家系间的变异分析结果同样可以看出,种源内家系间也存在丰富的变异,各生长阶段各性状种源内家系间平均变异系数均在0.10~0.38之间,15年生时,单株材积种源内家系间变异系数最大的是17233号种源,变异系数为0.60。比较各性状种源间与种源内家系间平均变异系数可以看出,各生长性状种源间的变异系数与种源内家系间平均变异系数相近,这说明黑荆树家系间的差异既有种源引起的效应也有种源内家系间的变异效应,因而开展种源选择与种源内家系选择对黑荆树改良都是有效的。
|
|

对试验林3,5,9,15年生时的观测数据进行种源及种源内家系2层次的方差分析结果(表 3)表明:种源及种源内家系各生长性状均达到极显著水平,但从方差分量上看,差异在种源水平上更大。3年生时,树高、胸径、树皮厚度及单株材积的种源方差分量分别是种源内家系方差分量的2~3倍;至15年生时,两者间的方差分量差距有所减小,树高、胸径、树皮厚度及单株材积的种源方差分量分别是家系方差分量的1.74, 1.35, 1.87和1.03倍。这一分析结果说明,黑荆树选育应从种源选择开始,在优良种源中选择优良家系能取得更好的选择效果。
|
|
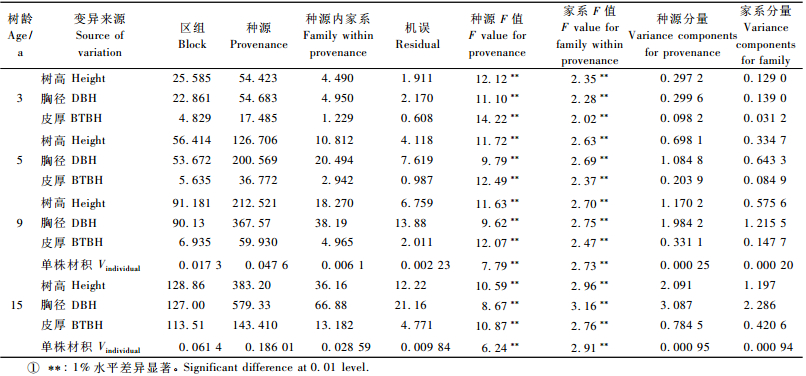
图 3为15年生时各种源树高、胸径、皮厚均值图,从图中可以看出,树高、胸径及树皮厚度在种源间差异较为明显。从树高、胸径两性状上看,17235,15858,14927三个种源生长量较为突出,其树高平均值在14 m左右,平均胸径生长量在13.7 cm以上; 而14769号种源表现最差,其树高、胸径平均值明显地低于其他种源。树皮厚度种源间的差异比起树高和胸径相对要小,树皮最厚的还是17235种源,平均厚度达到8.46 mm,其次是14925和14926,最薄的为14769号种源,平均厚度只有4.88 mm。

|
图 3 参试种源平均生长量 Figure 3 Trait means for provenances |
对参试家系15年生时生长性状均值的多重比较(LSD法)结果表明:树高显著高于总体均值的家系17个,其中极显著高于总体均值的家系有7个,分别是155,142,65,60,63,61,104号家系;胸径显著高于总体均值的家系有21个,极显著高于总体均值的家系10个,分别是142,63,61,155,60,158,74,36,104,132号;树皮厚度显著高于总体均值的家系有21个,极显著高于总体均值的家系15个,厚度从大到小依次为63,183,142,138,182,181,104,36,155,160,132,60,158,153,159号。单株材积显著高于总体均值的有7个,极显著高于总体均值的家系仅有1个,即63号家系。
2.3 遗传参数估算家系子代生长性状方差分量和遗传力反映了性状变异受遗传制约的程度,其大小直接影响选择的效果。黑荆树子代测定林不同生长阶段树高、胸径和树皮厚度的方差分量、单株和家系遗传力及遗传、表型变异系数见表 4。
|
|
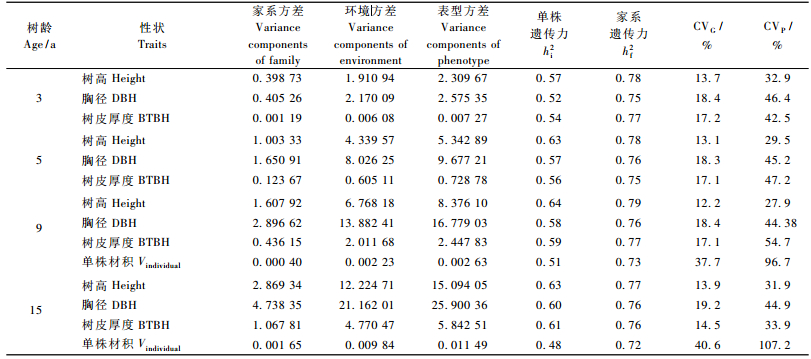
各性状的单株遗传力为0.54~0.57,家系遗传力为0.75~0.78,遗传力水平较高,说明黑荆树各生长性状受中等或中等以上遗传控制,具有很强的家系及家系内单株选择潜力。
遗传变异系数反映了遗传因素导致性状观测值相对变异的程度,黑荆树子代测定林直接观测性状遗传变异系数为12.2%~19.2%,胸径遗传变异系数最大,15年生时达到19.2%,其表型变异系数达到44.9%;而以树高、胸径估算的单株材积家系间的表型和遗传变异系数更是高达40.6%和107.2%,这说明黑荆树家系间存在丰富的变异。
2.4 生长性状年度相关与早期选择以3, 5, 15年生时黑荆树自由授粉家系子代测定林生长性状的观测数据为研究材料,对家系的各生长阶段生长性状进行协方差分析,并计算主要性状的早晚表型相关、遗传相关和环境相关系数,以了解黑荆树家系各生长性状的年度变异关系。表 5为参试家系早期(3, 5年生)胸径、树高和树皮厚度与生长后期(15年生)的树高、胸径、树皮厚度及单株材积间的表型、遗传和环境相关系数。
|
|

由表 5可以看出,黑荆树家系幼龄期(3, 5年生)的树高、胸径和树皮厚度观测值与成熟期(15年生)时树高、胸径和树皮厚度的观测值及单株材积计算值间的早晚表型及遗传相关系数随着树龄的增长而呈上升的趋势。3年生树高、胸径和皮厚与15年生时同一性状的早晚表型及遗传相关系数分别在0.65和0.75以上(P < 0.01),极显著相关。而5年生时各性状对15年生同一性状的表型及遗传相关系数已达到0.80和0.85以上,胸径的早晚表型及遗传相关系数更是高达0.86和0.97,相关非常紧密,说明5年生时的性状表现已完全能表达成熟林时的信息。
从早晚间接性状的相关系数也可看出,5年生时树高、胸径与15年生时单株材积间的表型及遗传相关系数均在0.78以上(P < 0.01),达到极显著相关。但比较树高和胸径对单株材积的相关系数可以看出,胸径对单株材积的年度相关系数明显大于树高对单株材积的相关系数,因此认为以胸径作为早期选择性状比树高更能表达成熟林的综合生长表现。
环境年度相关系数与表型及遗传相关系数的变化趋势一致,同样随着年龄的增长而递增,幼龄胸径对15年生胸径的相关系数由3年生时的0.58升至5年生时的0.83,胸径对单株材积的相关系数也由3年生时的0.49升到0.76,说明环境对树木生长表型有一定的影响。但比较环境与遗传的早-晚相关系数可知,幼年与15年生时生长性状的环境相关系数明显小于与之对应的遗传相关系数,说明黑荆树表型受遗传的影响大于环境影响。
生长性状早期选择遗传增益和相关遗传进度是确定早期选择年龄的主要依据,一般认为相关遗传进度最大、早期选择遗传增益稳定时的年龄即为最佳的早期选择年龄。表 6列出了3,5,9年生黑荆树树高、胸径和树皮厚度在入选率为20%(i=1.40) 时对15年生胸径、树高、树皮厚度及单株材积进行早期选择的遗传增益及相关遗传进度。括号内数据为根据相同性状进行早期选择的年选择效率,即早期选择的年遗传增益相对于成熟龄直接选择的年遗传增益的百分比。
|
|
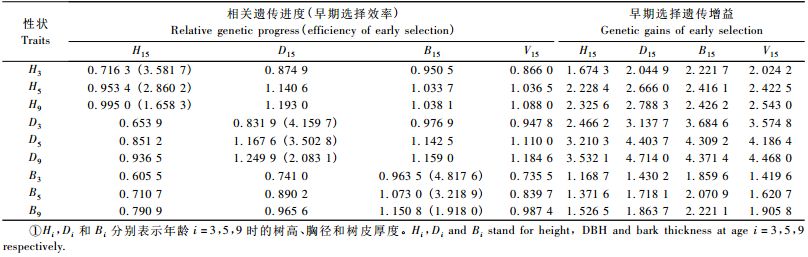
从表 6可以看出,各生长性状早-晚直接选择或间接选择的早期选择遗传增益及相关遗传进度均随树龄的增加而增大,而早期选择效率随树龄增加而下降。比较树高、胸径及树皮厚度直接选择的早期遗传增益和相关遗传进度发现,同一性状直接选择的早期遗传增益和相关遗传进度3年生明显小于5, 9年生,而5与9年生两者间的差异较小,说明5年生后遗传增益已接近最大值并趋向稳定,且此时也有较高年选择效率,而9年生时选择效率比5年生时有较大幅度的降低。综合早期遗传增益、相关遗传进度及早期选择效率的分析结果可以认为黑荆树家系早期选择在5~9年生间进行是比较合适的。另外,从幼龄性状对成熟林单株材积的间接结果可以看出,胸径对单株材积的早期选择增益及相关遗传进度均明显大于树高和树皮厚度,因而,胸径应是单株材积最可靠的早期间接选择性状。
3 结论与讨论 3.1 黑荆树生长性状变异受黑荆树生物学特性(雌雄同花,花量大,控制授粉困难;无性繁殖成活率不高等)限制,黑荆树的育种很难依靠杂交和无性系策略,因而种源和家系选择是黑荆树遗传改良的主要途径。黑荆树在澳大利亚自然分布区纵跨10个纬度带,横跨11个经度区,存在丰富的自然变异,国内外有关黑荆树种源及家系子代测定试验生长早期研究结果认为,黑荆树地理种源间在树高、胸径、干型等性状上均存在显著的差异(高传璧等,1994;张庆华等,1995;Searle et al., 2000),黑荆树生长早期家系间存在显著差异,主要生长性状遗传力较强(任华东等,1994)。商业栽培黑荆树的主要目的是生产木材和栲胶原料,即提高单位面积木材和树皮产量是黑荆树育种的主要目标,但与木材和树皮产量相关的性状是多方面的,主要是树高、胸径及树皮厚度(林杰等,1987)。本文对黑荆树166个自由授粉家系15年生子代测定林不同生长阶段的主要生长性状进行了种源/家系2层次变异分析,分析结果与前人的研究结果相似,黑荆树种源及家系间胸径、树高、树皮厚度、单株材积等生长性状存在着极显著的差异,这些差异主要来源于种源层次,同时种源内家系间也存在着丰富的变异,说明这些生长性状无论在种源水平上,还是家系水平上都存在着很大的改良潜力。
对黑荆树家系各主要生长性状的遗传变异分析表明,黑荆树单株和家系遗传力随树龄的波动不大,始终维持在较高的水平,受中等以上的遗传控制,说明黑荆树在其生长的不同阶段所受的遗传控制基本相同;在几个主要观测性状中表型及遗传变异系数胸径明显大于树高,说明胸径性状家系间变异更显著,这也从另一方面证明胸径是黑荆树种源/家系选择最有效的性状。
本文对种源/家系间的树皮厚度分析结果表明,树皮厚度性状在种源及家系间均存在极显著的差异,由于树皮厚度与树皮产量直接相关,如果选育目标是栲胶原料资源,那么在黑荆树种源或家系选择时,除考虑树高、胸径两性状外,树皮厚度也应作为重要指标,不能简单地用基于树高、胸径为自变量的二元公式(林杰等,1989)来估算单株树皮产量。
3.2 生长性状早晚相关性与早期选择获取单位时间内的最大遗传增益是林木遗传改良的目的,但由于林木生长周期长,林木遗传改良计划必须寻求一种可以缩短育种周期的方法,有关学者对杉木(Cunninghamia lanceolata)(阮梓材等,1999;马常耕等,2001)、马尾松(Pinus massoniana)(王章荣等,1987)、日本落叶松(Larix kaempferi)(丁振芳等,1997;孙晓梅等,2004; 2008)、西南桦(Betula alnoides)(郭文福等,2008)的研究结果认为林木早期选择是可行的,各树种在性状和适宜年龄上具有特异性。本文利用15年生黑荆树子代试验林3, 5, 9, 15年生时的观测材料,对幼、中龄生长性状与成熟龄(15年生)时的生长性状进行相关分析,结果表明不同阶段早-晚间均表现出不同程度的相关,自3年生始,树高、胸径和树皮厚度早-晚表型相关均已达显著水平,在5年生时各性状的早晚直接相关系数都在0.8以上,已达到极显著相关,遗传相关系数明显大于环境相关系数,说明黑荆树树高、胸径和树皮厚度早-晚间相关紧密,且这种相关关系主要受遗传因素控制,黑荆树早期选择具有较高的可靠性和有效性。
综合家系遗传力、相关系数、遗传增益和早期选择年效率的年度变化趋势,可以看出黑荆树5年生时树高、胸径、树皮厚度及单株材积的遗传力、相关系数及遗传增益均已趋于稳定,且年选择效率也处于较高水平,因而,5年生可以作为黑荆树家系选择的起始年龄。9年生时虽然与15年生时的早-晚相关系数最大,但此时的年选择效率比5年生时明显降低,因此,总体而言,从5年生开始至9年生前是黑荆树家系选择的合适年龄。另外,胸径早-晚间的遗传相关系数、早期选择增益及相关遗传进度均大于树高,与单株材积的间接相关系数也明显大于树高,同时随着年龄的增加树高的测量难度增大,测量误差增大,因此用胸径作为黑荆树早期选择指标较树高更合适。
3.3 黑荆树育种策略探讨从本文的分析结果可以看出,黑荆树家系间的差异更多源于种源层,因而在黑荆树的育种程序上,应先从种源试验开始,在种源试验基础上,从优良种源区选择优树并采集优树种子进行家系子代测定,将会取得更好的改良效果。本研究由于试材较多,出于田间试验环境控制需要,采用了单株小区,这种方式虽对黑荆树首次家系初步选择是可行的(Williams et al., 1994),但因小区数据缺失对研究结果产生了一定的影响,因而在开展进一步的家系选择时应尽可能采用多株小区。
丁振芳, 王景章. 1997. 日本落叶松家系早期选择技术[J]. 东北林业大学学报, 25(3): 65-67. |
高传璧, 郑芳楫, 任华东, 等. 1994. 黑荆树优良地理种源选择研究[J]. 林业科学研究, 7(2): 133-142. |
郭文福, 曾杰, 黎明. 2008. 广西凭祥西南桦种源家系选择试验Ⅰ.幼林生长性状的变异[J]. 林业科学研究, 21(5): 652-65. |
贺近格, BrownA G. 1991. 黑荆树及其利用[M]. 北京: 中国林业出版社: 5-320.
|
林杰, 陈平留, 黄健儿, 等. 1987. 黑荆树经营数表编制的研究(Ⅰ)立木材积表的编制[J]. 福建林学院学报, 7(2): 6-10. |
林杰, 陈平留, 黄健儿, 等. 1989. 黑荆树经营数表编制的研究(Ⅱ)经济材出材量表与树皮材积、重量表[J]. 福建林学院学报, 9(2): 127-133. |
马常耕, 周天相, 徐根良. 2001. 杉木无性系生长的遗传控制和早期选择初探[J]. 林业科学, 36(专刊1): 62-69. |
秦国峰, 周志春, 金国庆, 等. 1997. 马尾松天然林优树自由授粉家系生产力评价[J]. 林业科学研究, 10(5): 472-477. |
任华东, 高传璧, 郑芳楫, 等. 1994. 黑荆树自由授粉家系子代测定研究[J]. 林业科学研究, 7(1): 7-12. |
阮梓材, 胡德活, 王以珊, 等. 1999. 杉木家系物候型与早期选择[J]. 林业科学研究, 12(3): 291-298. |
孙晓梅, 张守攻, 侯义梅, 等. 2004. 短轮伐期日本落叶松家系生长性状遗传参数的变化[J]. 林业科学, 40(6): 68-74. DOI:10.11707/j.1001-7488.20040612 |
孙晓梅, 张守攻, 周德义, 等. 2008. 落叶松种间及种内和种间杂种家系间的物候变异与早期选择[J]. 林业科学, 44(1): 77-84. DOI:10.11707/j.1001-7488.20080113 |
王章荣, 陈天华. l987. 福建华安马尾松生长早晚期相关及早期选择[J]. 南京林业大学学报, 11(3): 41-47. |
张庆华, 张毅, 代宗贵, 等. 1995. 黑荆树地理种源选择增益研究[J]. 经济林研究, 13(3): 29-31. |
周志春, 林荣联, 兰永兆, 等. 1999. 马尾松实生种子园的遗传分析和育种值预测[J]. 林业科学研究, 12(2): 132-138. |
祝列克. 2001. 新世纪中国林木遗传育种发展战略[J]. 南京林业大学学报, 25(1): 3-8. |
Healy M J R, Westmacott M H. 1956. Missing values in experiments analysis on automatic computers[J]. Applied Statistics, 5: 203-206. DOI:10.2307/2985421 |
Payne R W, Lane P W, Ainsley A E, et al. 1989. Genstat 5 Reference Manual[M]. New York: Oxford University Press: 389-448.
|
Searle S D, Bell J C, Moran G F. 2000. Genetic diversity in natural populations of Acacia mearnsii[J]. Australian Journal of Botany, 48: 279-286. DOI:10.1071/BT98043 |
Snedecor G W, Cochran W G. 1980. Statistical Methods. 7th ed. Ames Iowa: Iowa State University Press, USA, 238-255. https://link.springer.com/chapter/10.1007/978-1-4613-2385-3_1
|
Williams E R, Matheson A C. 1994. Experimental design and analysis for tree improvement[J]. Melbourne, Australia: CSIRO Press: 93-116. |