 2010, Vol. 46
2010, Vol. 46文章信息
- 党宏忠, 张劲松, 赵雨森
- Dang Hongzhong, Zhang Jinsong, Zhao Yusen
- 应用热扩散技术对柠条锦鸡儿主根液流速率的研究
- Application of the Thermal Dissipation Probe Technique in Studying the Sap Flow in Taproot of Caragana korshinskii
- 林业科学, 2010, 46(3): 29-36.
- Scientia Silvae Sinicae, 2010, 46(3): 29-36.
-
文章历史
- 收稿日期:2008-11-04
-
作者相关文章


2. 东北林业大学 哈尔滨 150040
2. Northeast Forestry University Harbin 150040
TDP(thermal dissipation probe)技术自Granier发明以来,因其组成相对简单(双针)、测量精度和可信度高、相对成本低等优势,正越来越多地被应用到林木单株蒸腾的测定中来(Lu et al., 2004),成为目前用来推算林分尺度水分利用比较准确的方法之一。TDP技术要求探针的长度不能深入到心材部分,因此这一技术主要应用在胸径满足探针长度要求的乔木树种的茎干部位。虽然现在已经有了10 mm或更短的探针,但受成本等方面的限制,这一技术在主干、枝条较短细或分枝性强的灌木树种液流测定中还没有得到应用。柠条锦鸡儿(Caragana korshinskii)具有主根明显、分枝性强等生物学特征,是我国干旱半干旱区大部分生境条件下具有普适性的优良豆科固氮灌木树种。近年来,热技术被运用到柠条蒸腾耗水的测定与分析中来,已有研究分别利用热平衡技术(沈振西,2005)和热脉冲技术(Xia et al., 2008;夏永秋等,2008)测定柠条枝条的液流,并推算和分析单株的水分利用状况,但由于技术本身以及灌木树种生物学特性的限制,尤其是从样枝到单株尺度扩展中的误差等因素的存在,在一定程度上削弱了研究结果的可靠性。本研究将利用TDP技术把探针植入柠条锦鸡儿主根近地表根段从而测定单株的水分利用状况,较好地避开了这类树种分枝性强、枝条细难以直接用探针以及从样枝到单株尺度扩展等不利条件,同时降低了以往探针植入树干部位时辐射等环境因子对探针的影响,为分枝性强灌木树种整株液流的测定提供了一种新的方法。
1 材料与方法 1.1 材料的选择试验地位于半干旱区黄土丘陵沟壑第Ⅴ副区的甘肃省定西市石家岔流域,35°43′17.6″ N,104°39′14.0″ E,海拔2 218 m,年均降水量427 mm,年平均气温7 ℃,土壤为黄绵土,流域植被以20年生人工带状灌木篱柠条锦鸡儿林为主。试验材料选择3株生长旺盛的柠条锦鸡儿单株,以根径相对最大、冠幅较圆满的第Ⅱ号样株为例进行分析,该样株冠幅南北向为3.10 m,东西向为2.40 m,平均高2.15 m,主根直径3.25 cm,叶面积4.43 m2。
为了在不间断数据观测的同时求得样株边材面积,在以样株为中心半径为10 m的临时样地内选择6株根生长状态良好、冠幅相近的柠条锦鸡儿单株,挖取主根样段,立即用锋利的铡刀切出光滑、平整的截面,用游标卡尺量取各样段相互垂直方向的皮厚、韧皮部、边材、心材宽度,计算出各样段的平均边材宽度、边材面积。统计显示边材面积(S,cm2)与根径(D,cm)间的回归关系为S=4.681 4D-8.044 7,R2=0.97,并据此推算出样株Ⅱ根段的边材面积为7.17 cm2。
1.2 TDP的安装与测定采用按Granier原理设计的TDP-10 mm热电耦探针,探针直径1.2 mm,宽40 mm,电阻48 Ω,加热电压1.5 V。已有研究(Katherine et al., 2007;Rafael et al., 2007)证实了10 mm针用于林木液流测定的可靠性。安装时间为2008年3月21日,安装前先从地面近根茎处沿根向下挖开宽约30 cm的土槽,在距地面约20 cm深处挖开一空穴,使约20 cm的根段露出,刷掉表面土,用游标卡尺量取直径后用专用电钻在根表面垂直于根的走向钻孔(钻头直径与深度均与探针匹配),孔内涂适量凡士林后插入探针,用泡沫塑料纸紧紧包裹,在上端包裹接口缝隙处用黄油密封,避免渗入水分。安装完后加固空穴以免坍塌,并以水平放置的直径5 cm的PVC细管与外界相通,以排湿与防积水,空穴大小以TDP及包裹不与周围土壤直接接触为原则。采用220 V交流转接12 V铅酸蓄电瓶连续供电,采用英国SQ2020数采器采集数据,每5 min记录1次数据。液流通量(Fd,m3·m-2s-1)、液流速率(Fs,g·h-1)的计算采用Granier公式, ΔT, ΔTmax的计算均按照Granier(1987)方法确定,其中ΔTmax以7天为时长(Granier,1987;Lu et al., 2004)。
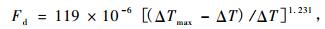
|
(1) |
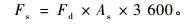
|
(2) |
式中: ΔTmax, ΔT分别为木质部液流为0和液流大于0时两探针间的温差(℃), As为加热探针处木质部的横截面积(m2)。
1.3 环境要素的测定与记录测定的环境要素类目及传感器类型见表 1,均与SQ2020数采器相连接,每5 min记录1次数据。
|
|

根据FAO Penman-Monteith方法(Allen et al., 1998), 在以天为步长计算潜在蒸发散ET0时,Cn=900,Cd=0.34;在以小时步长计算ET0时,Cn=37,Cd的取值根据文献(Allen et al., 2006)进行修正,其中白天(6: 00—20: 00)Cd=0.24,晚间(21: 00—5: 00)Cd=0.90。土壤热通量G的计算根据以小时为时间尺度计算时的精度要求,以固定系数乘以净辐射计算而得,该系数在白天、晚间分别取0.5和0.1(Allen et al., 1998)。
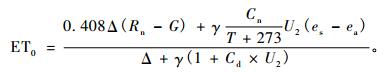
|
(3) |
式中:Rn为净辐射(MJ·m-2d-1或MJ·m-2h-1), G为土壤热通量(MJ·m-2d-1或MJ·m-2h-1),T为2 m高处温度(℃),U2为2 m高处风速(m·s-1), es, ea分别为饱和水汽压和实际水汽压(kPa),Δ为饱和水汽压曲线斜率(kPa·℃-1), γ为干湿表常数(kPa·℃-1),Cn, Cd是随参考作物类型和时间尺度而变化的常数。
2 结果与分析选择2008年4月30日—9月5日30天典型晴天日观测数据,在Origin 8.0程序下进行分析。
2.1 典型晴天日根部液流的日变化规律在众多环境要素中,太阳辐射(PY,W·m-2)、大气水分亏缺(VPD,kPa)是影响树干液流日变化的主导环境要素(Wullschleger et al., 1998;2001)。从图 1对柠条锦鸡儿根部液流的日变化中可以看出,典型晴天日PY呈典型的正态分布,从6: 00开始逐渐升高,13: 00达到峰值后逐渐下降,约20: 00趋于0,上升与下降时间段均约为7 h; 同样,根据太阳辐射、大气温度、大气湿度、风速、海拔、地理位置等综合要素计算的潜在蒸发散也表现出类似的日变化规律,13: 00的平均值约为(0.74±0.07)mm·h-1,最高值为0.87 mm; VPD并没有呈现出与PY, ET0一致的分布,而是呈偏峰的正态分布,始于6: 00的上升段并没有在PY最大时的13: 00达到峰值,而是延迟到约15: 00,之后又呈较快的下降趋势,夜间20: 00后呈缓慢下降趋势。

|
图 1 晴天时柠条锦鸡儿根部液流的日变化动态 Figure 1 Daily dynamics of sap flow in taproot of C. korshinskii in sunny days |
与环境要素的日变化趋势不同,液流分布呈宽峰的正态分布(图 1),并且可划分为phase 1~4阶段:phase 1阶段(6: 00—12: 00)液流呈快速上升;phase 2阶段(12: 00—16: 00)液流变化比较平稳,液流速率平均值能保持在97.41 g·h-1以上,峰值在13: 00为(99.32±19.36)g·h-1;phase 3阶段(16: 00—20: 00)液流又有一个比较快速的下降,但在光照约降为0后的20: 00时,液流速率仍能保持(18.40±6.89)g·h-1的水平;20: 00之后为phase 4阶段,液流缓慢下降,21: 00时也有平均值为(8.55±3.22)g·h-1的液流,整个夜晚液流平均值在3.62 g·h-1以上。液流的这种昼夜变化规律在日本柳杉(Cryptomeria japonica)等树种中也有相似报道(Takahisa et al., 2005)。
结合图 1对环境变量的日进程分析可以看出,在phase 1, phase 3阶段,太阳辐射是液流变化的最直接、最主要驱动力,但在VPD达到一定值后植物气孔调节行为启动,使得液流在phase 2阶段虽有较强的蒸腾需求拉力但液流变化缓慢。在phase 4阶段,柠条锦鸡儿呈现出明显的夜间液流活动,目前已有多篇文献报道了不同树种的枝干在夜间存在液流现象,并试图从夜间蒸腾(Green et al., 1989;Michael et al., 2006)、树干与叶片间运输距离延滞(Jan et al., 2007)以及补充组织失水(Senock et al., 1999;Joshua et al., 2007)等多方面给予解释,从对柠条锦鸡儿根部液流持续存在、冠高等综合因素来分析,更倾向于夜间液流用于补充白天旺盛的蒸腾带来的暂时的水分亏缺。统计表明,柠条锦鸡儿夜间液流量平均占全天的3.83%,低于纸桦(Betula papyrifera)的10%、红槲栎(Quercus rubra)的8%,但高于红花槭(Acer rubrum)的2%(Michael et al., 2006)。
2.2 根部液流与环境要素间的关系图 1展示了典型晴天日主要环境要素与根部液流的变化态势,为了进一步明确与液流变化间存在紧密关系的主要环境变量,对以上30天晴天日的6个主要环境要素与根部液流间做了相关分析,并以同期30天非典型晴天日(多云、阴天)为对照进行比较分析,各因子间的Pearson系数见表 2。
|
|
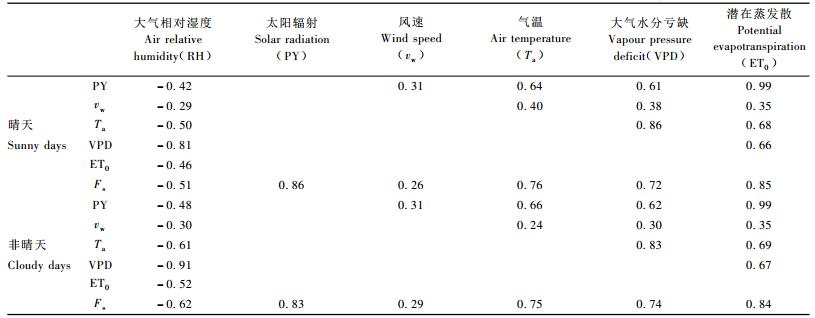
从表 2可以看出,在晴天日,与根部液流速率关系紧密的环境要素主要有PY(Pearson系数为0.86)、ET0(0.85),其次为Ta(0.76)和VPD (0.72)等; 在非晴天日,ET0(0.84)和PY(0.83)为主要环境要素,其次为Ta(0.75)和VPD(0.74)。可见,无论是晴天日还是非晴天日,太阳辐射是影响柠条锦鸡儿根部液流最主要的环境要素之一。有研究认为太阳辐射能解释树木白天液流变化方差的56%~71%,其他环境要素中以VPD为主(Thomas et al., 2005)。多元回归表明,Fs与各环境要素间具有以下回归关系:
晴天日:Fs=0.248 PY + 0.641 Ta + 17.841 VPD -262.561 ET0, R2=0.92;
非晴天日:Fs=0.186 PY + 0.423 Ta + 22.20 VPD -152.411 ET0, R2=0.88。
Fs与PY和ET0的回归关系式分别为:
晴天:Fs=0.038 PY + 107.908 ET0, R2=0.85;
非晴天:Fs=-0.050 PY + 261.372 ET0, R2=0.82。
太阳辐射与ET0能解释液流速率80%以上的变差,与只考虑PY与VPD 2个变量相比(Wullschleger et al., 1998;2001),更为可靠的是预估液流速率的环境变量。ET0综合了包括辐射在内的环境因素如VPD, vw, Ta等,因而能更全面地反映大气蒸发潜力。有研究也表明在众多气候变量中,ET0与树木液流间具有更为密切的关系(Conejero et al., 2007),Granier(1987)对花旗松(Pseudotsuga menziesii)液流的研究认为这种关系是线性(R2=0.94),本次研究也例证了ET0与根部液流间具有较紧密的相关关系(图 1, 表 1),可以认为它是分析根部液流理想的环境变量。
2.3 不同时段根部液流变化与环境因子间的响应关系 2.3.1 太阳辐射如前所述,柠条锦鸡儿根部液流主要集中在有光照的白天,白天大气巨大的蒸腾驱动力促使根系从土壤中源源不断地吸收水分,从而保证了叶片等器官组织的水分需求。但是液流随光照的变化在上午和下午表现出不同的变化规律(图 2a箭头方向所示),上午,随着光照强度的增加,液流随光照增加而升高的趋势基本呈线性(图 2b),其拟合方程:Fs=0.107 8PY(R2=0.95),约13: 00时光照、液流均达到最高(这与图 1所示相同),这表明试验期间PY是液流增加的主要驱动力,尚不足以成为限制液流增加、诱导植物气孔调节行为发生的主要因素。下午,随着PY的下降,Fs下降相对缓慢,在光照下降到200 W·m-2左右时,Fs才有较快的下降(图 2c),用Sigmoidal-Hill函数拟合的方程为:Fs=151.15 PY0.454/(161.700.454+PY0.454)(R2=0.84)。由于光合总是伴随着水分的蒸腾,因此这可能进一步揭示柠条锦鸡儿叶片气孔充分利用光照积累干物质的特性。

|
图 2 不同时段太阳辐射对液流速率的影响 Figure 2 Effects of solar radiation to sap flows in different time of the day |
ET0具有与PY相似的日变化规律(图 1),因此,柠条锦鸡儿根部液流随ET0的变化也呈现出与PY相近的变化趋势,尽管有报道建立ET0与Fs间的线性关系(Antonio et al., 2006;Xia et al., 2008),但从图 3a中可以看出, 在日变化中Fs与ET0间并不是单一的线性关系。上午,随着ET0的增加,液流呈近似线性增加(图 3b),拟合方程:Fs=150.85 ET0(R2=0.94),约在13: 00时ET0, Fs均达到最高,之后,Fs随ET0的降低而缓慢下降,在ET0下降到约0.3 mm·h-1后,Fs快速下降(图 3c),用Sigmoidal-Hill函数拟合的方程为:Fs=105.23 ET01.66/(0.0861.66+ ET01.66)(R2=0.74)。这说明试验期间大气蒸发潜力并没有成为限制液流的因素,可能与柠条锦鸡儿属于主根垂直分布、深根性树种有关,土壤水分对液流日变化的影响没有大气环境要素表现得明显。

|
图 3 不同时段ET0对液流速率的影响 Figure 3 Effects of ET0 to sap flows in different time of the day |
对于图 1展示的Fs, VPD的日变化趋势,有必要进一步明确其相互关系,为此对不同时段Fs随VPD的变化进行了分析。从图 4a中可以看出,上午,Fs随着VPD的增加而快速升高(排除因土壤水分严重不足影响液流的可能性),在VPD约高于1.11 kPa[约10: 00,Thomas等(2005)认为这一值为1~1.2 kPa]后,Fs增幅变缓并在VPD约为2 kPa时达到峰值(99.32 g·h-1,约在13: 00),之后维持在较高水平直到VPD于15: 00达到峰值(2.35 kPa)后(此时Fs约为97.41 g·h-1),Fs随VPD的下降而快速下降。从图 4a中还可以看出,与PY, ET0对Fs的影响不同,在同一VPD值时,下降阶段(对应时段为15: 00—20: 00)的Fs值明显低于上升阶段(对应时段为6: 00—15: 00)的Fs值(图 4a箭头所示方向),这种规律在对红花槭等树种的研究中也有报道(Wullschleger et al., 1998),这与树木在午间受到较强的水分胁迫后,通过气孔调节来维持组织膨压的反应有关,土壤水分环境越好,这种规律越趋于明显(Wullschleger et al., 1998)。从图 4b(Sigmoidal-Hill拟合方程:Fs=100.246VPD3.95/(0.6783.95+ VPD3.95)(R2=0.73)可以看出,2 kPa是柠条锦鸡儿Fs随VPD变化由上升转为下降的拐点(约在13: 00),这可能也是柠条锦鸡儿气孔调节行为发生、液流受抑制时的VPD阀值,这一值与Ortuno等(2006)对柠檬(Citrus limonum)等树的研究相近。同时结合图 4c(线性拟合方程:Fs=36.328 VPD,R2=0.87)可以看出,气孔调节对液流变化影响更明显的拐点应该是VPD平均值约为2.35 kPa(约在15: 00),此后,Fs随VPD的减弱呈快速下降趋势。Hogg等(1997)研究表明即便在VPD维持较高水平时,液流在光照低于400 W·m-2时也开始迅速下降,佐证了太阳辐射对气孔行为的重要影响。结合本研究中对Fs受PY(图 2)、ET0(图 3)的分析认为,柠条锦鸡儿因较高的VPD诱导的气孔行为对液流的调控作用要比因大气蒸发需求下降的影响更加明显,由于柠条锦鸡儿是深根性树种以及根际土壤水分变化缓慢,更加相信这可能是柠条锦鸡儿叶片应对午间水分亏缺的主要生存策略。

|
图 4 不同时段大气水分亏缺对液流速率的影响 Figure 4 Effects of VPD to sap flows in different time of the day |
柠条锦鸡儿根部液流在晴天日表现出典型的宽峰型正态分布,并可划分为phase 1~4阶段, 不同阶段的差异揭示了太阳辐射、大气水分亏缺等大气环境因子对液流的直接驱动效应,以及柠条锦鸡儿自身叶片气孔行为对液流变化的调控作用,表明环境与树木自身共同影响液流变化。大气水分亏缺是启动气孔调节行为的最主要环境因子。在众多环境因子中,ET0综合性强且与树木液流速率相关系数较高,是估算液流更为可靠、理想的环境变量。柠条锦鸡儿夜间持续存在液流现象,平均值约在3.62 g·h-1以上,但夜间液流所占全天比例仅为3.83 %,这也是对Joshua等(2007)关于绝大部分植物夜间都有蒸腾液流的一个例证。白天液流速率随环境因子的变化在不同时段表现出不同的规律,在早晨启动的上升阶段, Fs与PY,ET0间可以用线性方程取得较好的拟合,峰值后的下降阶段用函数Sigmoidal-Hill拟合较好,而Fs与VPD间的拟合关系与此相反。
图 2,3中Fs随PY, ET0上升阶段的近线性应该与植物叶片进行光合作用时的主动性有关(必然伴随水分消耗过程),足够的光照是激活叶绿素及系列光合过程链条的前提,足够的大气蒸发需求也是形成大气-叶-根足够水分压力梯度、促使木质部导管水分上升的前提,因而这一阶段PY, ET0均是表达液流活动驱动力的主要变量; 而下降阶段呈Sigmoidal-Hill函数关系(先缓慢后快速下降)也恰恰说明植物关闭部分或全部气孔行为的被动性,这一点也能从图 4中VPD与Fs的变化曲线中得到例证。事实上图 4a中从10: 00 (VPD约为1.11 kPa)开始Fs增幅变缓慢已表明VPD是液流活动中的主要阻力变量,15: 00后Fs才开始下降(而且是较陡然的近线性下降)也说明气孔活动与大气水分亏缺间的“斗争”,VPD启动的气孔行为并进而产生的液流速率降低对植物来说并不一定是坏事。
由于树干的比输导率(单位时间通过一定长度和横切面茎段移动水分的容积)是从基部向顶端减小,因此高大乔木树种可能在根系吸水、树干液流及叶片蒸腾间存在时滞,测定部位对于准确估计液流非常关键,本研究选择在主根近地表根段安装热电藕探针,保证了所监测液流为单株液流。从本次研究来看,柠条锦鸡儿根部液流随主要环境要素的变化在时间上是近似同步的,没有出现典型的时滞现象,这一方面是由于灌木树种树高相对较低,另一方面可能是由于木质部中纵向水流的阻力在根中要比茎中低很多(Kramer et al., 1979),因此在根部测定液流并用来计算灌木树种单株耗水的方法具有明确的科学性与优越性。尽管本试验中柠条锦鸡儿样株主根直径只有3.25 cm,液流的径向差异应该较小,但随着试验技术的改进,这一判断将得到进一步的验证。
夏永秋, 邵明安. 2008. 黄土高原半干旱区柠条(Caragana korshinskii)树干液流动态及其影响因子[J]. 生态学报, 28(4): 1376-1382. |
沈振西. 2005. 宁夏南部柠条、沙棘和华北落叶松的液流与蒸腾耗水特性. 中国林业科学研究院博士后研究工作报告.
|
Allen R G, Pereira L S, Raes D, et al. 1998. Crop evapotranspiration: Guidelines for computing crop water requirements[J]. FAO Irrigation and Drainage Paper, 56: 301. |
Allen R G, William O P, James L W, et al. 2006. A recommendation on standardized surface resistance for hourly calculation of reference ET0 by the FAO 56 Penman-Monteith method[J]. Agricultural Water Management, 81: 1-22. DOI:10.1016/j.agwat.2005.03.007 |
Antonio R P, Steve G N, Augusto V N. 2006. Penman-Monteith reference evapotranspiration adapted to estimate irrigated tree transpiration[J]. Agricultural Water Management, 83: 153-161. DOI:10.1016/j.agwat.2005.11.004 |
Conejero W, Alarcon J J, Garcia O Y, et al. 2007. Daily sap flow and maximum daily trunk shrinkage measurements for diagnosing water stress in early maturing peach trees during the post-harvest period[J]. Tree Physiology, 27: 81-88. DOI:10.1093/treephys/27.1.81 |
Granier A. 1987. Evaluation of transpiration in a Douglas fir stand by means of sap flow measurements[J]. Tree Physiol, 3: 309-320. DOI:10.1093/treephys/3.4.309 |
Green S R, McNaughton K G, Clothier B E. 1989. Observations of night-time water use in kiwifruit vines and apple trees[J]. Agric For Meteorol, 48: 251-261. DOI:10.1016/0168-1923(89)90072-5 |
Hogg E H, Hurdle P A. 1997. Sap flow in trembling aspen: implications for stomatal responses to vapor pressure deficit[J]. Tree Physiology, 17: 501-509. DOI:10.1093/treephys/17.8-9.501 |
Jan C, William L B, Nathan P, et al. 2007. Tree water storage and its diurnal dynamics related to sap flow and changes in stem volume in old-growth Douglas fir trees[J]. Tree Physiology, 27: 181-198. DOI:10.1093/treephys/27.2.181 |
Joshua B, Fisher, Dennis D, et al. 2007. What the towers don't see at night: nocturnal sap flow in trees and shrubs at two Ameri Flux sites in California[J]. Tree Physiology, 27: 597-610. DOI:10.1093/treephys/27.4.597 |
Katherine A, Mcculloh, Klaus W, et al. 2007. A comparison of daily water use estimates derived from constant-heat sap-flow probe values and gravimetric measurements in pot-grown saplings[J]. Tree Physiology, 27: 1355-1360. DOI:10.1093/treephys/27.9.1355 |
Kramer P J, Kozlowski T T. 1979. Physiology of woody plants[M]. New York: Academic Press.
|
Lu P, Urban L, Zhao P. 2004. Granier's thermal dissipation probe(TDP) method for measuring sap flow in trees: theory and practice[J]. Acta Botanica Sinica, 46(6): 631-646. |
Michael J D, Nathan G P. 2006. Interspecific variation in nighttime transpiration and stomatal conductance in a mixed New England deciduous forest[J]. Tree Physiology, 26: 411-419. DOI:10.1093/treephys/26.4.411 |
Ortuno M F, Garcia O Y, Conejero W, et al. 2006. Relationships between climatic variables and sap flow, stem water potential and maximum daily trunk shrinkage in lemon trees[J]. Plant and Soil, 279: 229-242. DOI:10.1007/s11104-005-1302-z |
Rafael P, Jan C, Pilar L. 2007. Variation in the radial patterns of sap flux density in pubescent oak(Quercus pubescens) and its implications for tree and stand transpiration measurements[J]. Tree Physiology, 27: 537-548. DOI:10.1093/treephys/27.4.537 |
Senock R S, Leuschner C. 1999. Axial water flux dynamics in small diameter roots of a fast growing tropical tree[J]. Plant and Soil, 208: 57-71. DOI:10.1023/A:1004494432610 |
Takahisa N, Hisashi A, Takeshi M, et al. 2005. The relationship between sap flow rate and diurnal change of tangential strain on inner bark in Cryptomeria japonica saplings[J]. J Wood Sci, 51: 441-447. DOI:10.1007/s10086-004-0686-3 |
Thomas M, Nicole M, Manfred K, et al. 2005. Stomatal conductance, transpiration and sap flow of tropical montane rain forest trees in the southern Ecuadorian Andes[J]. Tree Physiology, 25: 1283-1293. DOI:10.1093/treephys/25.10.1283 |
Wullschleger S D, Hanson P J, Todd D E. 2001. Transpiration from a multi-species deciduous forest as estimated by xylem sap flow techniques[J]. Forest Ecology and Management, 143: 205-213. DOI:10.1016/S0378-1127(00)00518-1 |
Wullschleger S D, Hanson P J, Tschaplinski T J. 1998. Whole-plant water flux in understory red maple exposed to altered precipitation regimes[J]. Tree Physiology, 18: 71-79. DOI:10.1093/treephys/18.2.71 |
Xia G M, Kang S Z, Li F S, et al. 2008. Diurnal and seasonal variations of sap flow of Caragana korshinskii in the arid desert region of north-west China[J]. Hydrological Processes, 22: 1197-1205. DOI:10.1002/(ISSN)1099-1085 |