 2010, Vol. 46
2010, Vol. 46文章信息
- 邓青珊, 陈思静, 鲁长虎
- Deng Qingshan, Chen Sijing, Lu Changhu
- 鸟类对南方红豆杉种子的取食与搬运
- Foraging and Transporting of Chinese Yew Seed by Frugivorous Birds
- 林业科学, 2010, 46(2): 157-161.
- Scientia Silvae Sinicae, 2010, 46(2): 157-161.
-
文章历史
- 收稿日期:2008-08-12
-
作者相关文章



种子植物天然更新过程大致包括种子生产、种子扩散和种子萌发生长等3个阶段,各阶段所涉及的生物和非生物因子很多(Nathan et al., 2000)。Chambers(1994)就种子扩散的动态过程提出了种子命运(seed fate)模型,将种子命运划分为2个阶段:第1扩散阶段,即可萌发的种子从植株(母树)到靠近母树的地面;第2扩散阶段,即种子在地面水平或地面到土层间垂直运动过程。第1扩散阶段中,种子依靠风力、重力等非生物因素离开母树到地面最为常见;然而,生物因子的影响也是一个非常重要的因素。已有的研究充分显示:被子植物中的肉质果(核果、浆果等)往往在成熟后被动物捕食(Johnson et al., 1985; 鲁长虎等, 1997)等,这些动物捕食者可能只消化果肉部分,而成为种子的扩散者(Murray, 1986; Levey et al., 2002; Forget et al., 2004)。
红豆杉属(Taxus)植物种子是裸子植物中独特的一类,种子被粘性较强的肉质假种皮紧紧包裹着,不易脱落,与被子植物肉质果实类似(为叙述方便,本文将假种皮完整的种子称为果实)。在种子成熟后,假种皮变红色且味甘甜,常常吸引鸟类和小型哺乳动物取食(DiFazio et al., 1996; Thomas et al., 2003)。相关研究表明,鸟类取食是欧洲红豆杉(T. bacatta)种子消失的主要途径(Hulme, 1996),如乌鸫(Turdus merula)、环颈鸫(Turdus torquatus)、槲鸫(Turdus viscivorus)等鸟类。落到地面的种子会被鼠类取食,如小林姬鼠(Apodemus sylvaticus)等(García et al., 2000)。在波兰的Kórnik植物园,普通鳾(Sitta europea)和画眉(Garrulax canorus)是主要的取食和传播动物。普通鳾偏爱停留在灌木和有着光滑树皮的树种上, 如山毛榉(Fagus spp.)、角树(Carpinus spp.),在这些树下多见红豆杉幼苗;画眉则可将红豆杉种子传播至更远处(Giertych, 2000)。由于鸟类只消化了种子外被的假种皮,种子可随粪便排出或被呕出,因此鸟类通过取食参与了欧洲红豆杉的天然更新过程。
南方红豆杉(T. chinensis var. mairei)是我国特有种、国家一级重点保护植物。自然条件下南方红豆杉常见分布在一些村落附近,由较古老的个体组成,数量一般在几十株左右,种群呈现濒危、衰退状态(费永俊等, 1997)。其天然更新主要通过种子繁殖实现。一些报道认为, 种子结实量少、动物取食种子、种子萌发难等是其更新不良的原因(费永俊等, 1997; 朱念德等, 1999)。本研究选取皖南仙寓山的南方红豆杉种群作为对象,于2006年和2007年研究了鸟类取食与搬运对种子命运的影响。
1 研究地自然概况仙寓山位于皖南石台、祁门、东至3县交界处,东南部为黄山北行山脉盘结,西北部是九华山脉的南端,形成南北高,东西低的地势。境内山峦起伏,沟壑纵横,海拔一般在50~1 000 m之间,最高海拔为1 375.7 m,森林覆盖率达90%,总面积51万hm2。属于中亚热带季风区,年平均气温16.1 ℃,年平均降雨量1 649.5 mm。
仙寓山地区南方红豆杉天然种群多零星分布在海拔540 m以上的陡坡地带。研究地点选择分布最为集中的石台县珂田乡双坑村附近。村庄位于谷底自西向东流淌溪涧的两侧(图 1),其北面山坡下坡是毛竹(Phyllostachys edulis)林,上坡是杉木(Cunninghamia lanceolata)林;南面山坡为红豆杉主要分布的坡面,该坡面的中间有一片地势低洼的茶园,茶园的东面为杉木林,西面为毛竹林。除茶园外,南北山坡上均分布一定数量的南方红豆杉幼树和幼苗。

|
图 1 仙寓山南方红豆杉研究区域示意图 Figure 1 Map of the distribution of T. chinensis var. mairei in study site in Xianyu Mt. ①~⑤: 1~5号树, 下同。 No.1-No.5 parent tree, the same below. |
成年个体零星分布在村庄南面山坡毛竹林(即西坡竹林)内,研究期间具结实能力的母树仅有5株(图 1),其中最大的一株树龄约145年,树高20 m以上,胸径达44.6 cm,冠幅达5 m(①号)。在其相距3~4 m处有2株结实量较少的母树,其中②号树树高为4.4 m,胸径为11.0 cm,③号树树高7.4 m,胸径13.5 cm。这3株母树位于坡地林缘。另2株母树相距较远,一株在①号树东南侧约20 m的中坡凸起的山崖后(④号),树高7.5 m,胸径12.4 cm,结实量一般;另一株在①号树西南侧约100 m的中坡上,树高13 m,胸径17.5 cm(⑤号),结实量较大。
2 研究方法 2.1 种子雨的确定分别于2006, 2007年10月—12月,观察记录5株南方红豆杉母树的种子雨持续时间。在本研究中种子雨特指南方红豆杉成熟的种子向环境中扩散的过程。以假种皮变红色作为种子成熟的标记,一株母树上开始出现红色果实视为种子雨开始,全部果实消失视为种子雨结束;全部5株种子雨时长作为种群种子雨的持续时间。
2.2 落果测定以①号母树为重点,在种子开始成熟后,进行测定。对每天新增的落果进行插桩标记,记录每天的落果量,并注意天气、鸟类取食等因素与落果数量的变化。观察落果的变化,统计每天果实腐烂、被昆虫取食情况以及种子的消失量。
2.3 鸟类取食和搬运观察以①号母树为重点,从种子雨来临开始,选择视野开阔,不影响鸟类等动物取食的地点为观察点,以10×26倍变焦双筒望远镜全天候(天亮至天黑,约6:30—17:00)观察,记录访问鸟类的种类、数量、取食行为、停留时间、取食量,全部观察直至树上无挂果、种子雨结束为止。2007年研究期间,在附近布置鸟网捕捉鸟类,进行人工饲喂,初步确定红豆杉种子经过消化道的时间。
3 结果与分析 3.1 南方红豆杉种子雨种子成熟的具体时间因立地条件和母树自身的生长状况而有所差异。研究期间5株母树的种子雨持续时间如下(表 1)。
|
|

① 号母树在2006年10月20日进入果熟期,10月25日后大量成熟,到11月19日后基本无挂果,种子雨持续时间为31天。2007年10月26日进入了果熟期,10月31日后大量成熟,到11月28日后基本无挂果,种子雨持续时间为34天。其余4株母树的种子雨开始时间在2006年比①号母树稍晚5~7天,2007年则接近。④号母树种子量在2年里均少,因而种子雨持续时间短。2007年⑤号母树因人为采果干扰,无法准确统计种子雨持续时间。总体上看,2006年全部种群的种子雨持续时间为38天,而2007年为34天,种子在母树上的存续时间短,消失的速度较快。
种子成熟后主要有2个途径从树上消失:一是直接掉落,二是鸟类直接从树上取食。由于在村落附近,人为采摘种子也是种子消失的途径之一,②、③和⑤号母树在2007年均有人为采果。
采用标准枝法,统计出2007年①号母树果实产量约为6 574颗。2007年10月26日至11月28日,掉落的落果累计有2 082颗,约占总数量的31.7%,均分布在母树冠幅下近20 m2范围内。
雨天对落果的影响大,11月1日、11月16日至11月18日均为大雨天气,新增落果量相应增多(图 2)。鸟类取食时振落是种子落地数量增加的另一重要因素。刚开始进入果熟期时,鸟类访问较少,落果多是自然脱落。比较新增落果和鸟类取食的趋势(图 2),在11月1日、11月9日和11月16日鸟类取食次数出现峰值,新增落果量相应增加。

|
图 2 2007年①号母树每日新增落果与鸟类取食次数 Figure 2 Number of increased fallen fruits of tree ① and foraging times of birds per day in 2007 |
1) 取食鸟类种类 研究期间共记录到12种鸟类从①号母树上直接取食果实,均为雀形目鸟类。7种体型较大的鸟采取整吞方式取食(表 2),即将果实从小枝上啄取后,整个吞下。另有5种体型较小到鸟类采取啄食方式,即直接在空中悬停啄食假种皮或将果实衔至附近再撕食假种皮,并不吞下种子,分别是红胁蓝尾鸲(Tarsiger cyanurus)、棕脸鹟莺(Abroscopus albogularis)、红尾水鸲(Rhyacornis fuliginosus)、灰眶雀鹛(Alcippe morrisonia)、红头长尾山雀(Aegithalos concinnus)。啄食取食虽然偶尔也搬运种子,但次数很少,因此只有整吞取食对种子搬运具有实际意义。
|
|
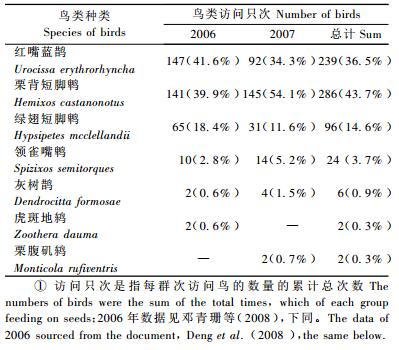
2006年记录到367取食只次,2007年为288只次,共计655只次。由于2年的观察周期相近,可以推测取食次数的减少与种子数量少有关(2007年结实量少于2006年)。从鸟类取食次数统计看,红嘴蓝鹊、栗背短脚鹎和绿翅短脚鹎为主要取食鸟类。2006年3种鸟类共计取食353只次,占当年鸟类取食总次数的96.2%;2007年共计268只次,占当年鸟类取食总次数的93.1%;2年总体上占94.8%。
红嘴蓝鹊在2年间取食差异较大。2006年红嘴蓝鹊几乎每天都会取食,2007年集中在雨水、大雾、大风天气取食,在晴好天气取食的仅记录有19只次。这与2007年人类活动的影响增大有关: ①号母树靠近村庄, 晴好天气时村民砍伐毛竹,而红嘴蓝鹊胆小,较栗背短脚鹎等鹎类更容易受惊。绿翅短脚鹎多是成对来取食,2006年发现其在坡上竹林中的巢,2007年则未见,其取食次数减少可能与此有关。栗背短脚鹎2年记录的取食次数基本相近。
2) 鸟类取食行为 红嘴蓝鹊多4~6只集群取食(表 3),最多时达每群10只,单只取食情况较少,仅6次。栗背短脚鹎多2~3只集群取食,最多时每群7~8只。绿翅短脚鹎最为稳定,多2只飞来取食,单只取食次数仅4次。在停留时间上,红嘴蓝鹊的平均停留时间较短,2种鹎类的停留时间均达数分钟。其中绿翅短脚鹎最长,2007年的记录为平均992.3 s,最长的1次达47 min,一般取食4~5颗后便停歇于树上相互鸣叫,数分钟或十几分钟后再进行1次取食。
|
|
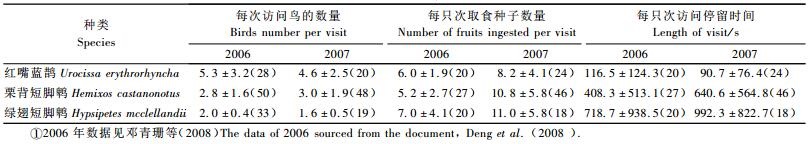
从单次访问取食量上看,2006年和2007年均以绿翅短脚鹎的取食量稍大,经单因素方差分析(one-way ANOVA)表明,3种鸟的取食量差异并不显著(2006年: F=2.001<F2, 72, 0.05=8.432;2007年: F=2.189<F2, 85, 0.05=3.118)。总体来说,2007年的取食量均比2006年的有所增加,增幅最多的为栗背短脚鹎,平均增加了1倍。
如果按单次取食量平均值计算,2006年3种主要鸟类取食①号母树种子约2 058颗,其中红嘴蓝鹊取食870颗,栗背短脚鹎733颗,绿翅短脚鹎455颗;2007年3种主要鸟类取食①号母树种子共约2 661颗,占①号母树果实产量的40.47%,其中栗背短脚鹎取食1 566颗,红嘴蓝鹊754颗,绿翅短脚鹎341颗。这进一步说明红嘴蓝鹊、栗背短脚鹎和绿翅短脚鹎是主要的取食鸟类。
3) 鸟类的搬运路径 鸟类取食后,种子经过消化道被搬运。搬运路径决定了南方红豆杉种子将被扩散多远及散落在何种生境中,3种主要取食鸟类的取食搬运路径有明显差异(图 3,参见图 1)。

|
图 3 2006年和2007年3种主要鸟类取食路径 Figure 3 The route of three species of birds feeding on T. chinensis var. mairei |
红嘴蓝鹊活动范围较大,在东、西、南、北4个方向都有一定数量的飞来, 飞离, 或沿溪向上游(西)、向下游(东)或飞至对面山坡(北)、或飞往南坡活动稍后再离开。以在沿溪向下(东)方向上活动最为频繁。栗背短脚鹎的活动范围基本上只限于南坡竹林内。取食完毕后或飞往东南方向①号树右侧约20 m的山崖后,有时至④号树上取食,或飞往西南方向①号树左侧约30 m的山崖后,有时沿竹林活动至⑤号树取食,或飞向①号母树正南坡。其中以飞往东南方向的活动频率最高。绿翅短脚鹎活动范围和路径与栗背短脚鹎的大致一样,基本上只在南坡竹林内活动。在东南方向①号树右侧约20 m的山崖的飞入和飞出频率最高。
4 讨论 4.1 种子从母树上消失的特点南方红豆杉种子成熟后,其假种皮变红而甘甜。此后,种子经历2个途径:或者自然掉落地面,或者被鸟类从树上直接取食。种子雨持续时间短,2年的记录可见最长的为34天(表 1),结实量少的母树则最短在17天内全部从树上消失,种子消失的速度快。由于1株母树上的种子是逐步成熟的,所以完全成熟的种子在树上的存续时间实际上更短,可能仅几天。
本研究中以2007年①号母树所做的统计表明:全部约6 574颗种子中的2 082颗直接掉落地面,占全部种子的31.7%。从2006年研究时在①号母树下发现的很多幼苗看(邓青珊等,2008),掉落地面的种子有一部分能够顺利萌发。而其他大部分种子则直接被从树上取食搬运,记录到白天3种主要鸟类平均取食搬运约2 661颗。相差的部分可以确定也被鸟类取食搬运,或者被夜间活动的啮齿类等动物搬运取食,在母树附近捕获的隐纹花鼠(Tamiops swinhoei)和黄鼬(Mustela sibirica),是否上树取食有待进一步确认。由此看来,①号母树全部种子约1/3直接掉落地面,约2/3主要被鸟类直接取食搬运离开母树。研究期间排除人为采摘种子外,其他母树鸟类取食搬运的情况与①号母树相似。
4.2 鸟类搬运种子的意义红豆杉种子成熟后不能单独脱落,而是随假种皮掉落母树下,且种子较大而不能随风扩散(朱琼琼等,2007)。鸟类将南方红豆杉的果实整吞后,果肉被消化,种子随粪便排出体外,粪便随机落在林内。本研究中在鸟类经常活动的地点发现带有种子的新鲜鸟类粪便,每个含有1~2颗种子。由于鸟类搬运的数量较大,在鸟类常活动的地方则会形成一个种子库。由于红豆杉种子需经历2个冬天后才能萌发(谭一凡,1991),种子库的种子将会受到啮齿类等动物的捕食,可以确信的是部分种子到达适宜萌发的小生境,并顺利萌发成为幼苗(邓青珊等,2008)。
本研究发现有7种鸟类通过整吞方式取食南方红豆杉果实,都具有扩散种子的作用。其中领雀嘴鹎、灰树鹊、栗腹矶鸫和虎斑地鸫4种食果鸟类的取食次数和取食量相对来说较少(表 2),对果实的扩散作用小。其余3种鸟类红嘴蓝鹊、栗背短脚鹎和绿翅短脚鹎对南方红豆杉具有重要的扩散作用。
栗背短脚鹎和绿翅短脚鹎取食红豆杉种子后,停留时间较长,且会多次取食或在相邻的树间取食,飞往途径和活动范围仅限于西坡竹林和东坡的杉木林(图 3和图 1)。根据野外捕获栗背短脚鹎喂食试验,消化道中的滞留时间最短仅为8 min,可以看出,经过栗背短脚鹎和绿翅短脚鹎取食和搬运后的红豆杉种子可能集中在离母树不远的竹林和杉木林坡面上,不做较远距离的搬运。红嘴蓝鹊同样也为集群活动取食,但取食种子后停留时间较短,取食完毕后会飞至较远安全处(图 3),因此经过红嘴蓝鹊取食和搬运后的红豆杉种子被移动至离母树较远的地方,对距离母树300~400 m北坡林下的红豆杉更新贡献较大(图 1)。从动植物相互作用角度看,鸟类获得了红豆杉提供的营养(假种皮),红豆杉种子获得了扩散的机会,鸟类与南方红豆杉之间形成了互惠(mutualism)关系(Levey et al., 2002)。
邓青珊, 朱琼琼, 鲁长虎. 2008. 南方红豆杉的天然更新格局及食果鸟类对其种子的传播[J]. 生态学杂志, 27(5): 712-717. |
费永俊, 雷泽湘, 余昌均, 等. 1997. 中国红豆杉属植物的濒危原因及可持续利用对策[J]. 自然资源, (5): 59-63. |
鲁长虎, 常家传. 1998. 食肉质果鸟对种子的传播作用[J]. 生态学杂志, 17(1): 61-64. |
谭一凡. 1991. 南方红豆杉种子后熟生理的研究[J]. 中南林学院学报, 11(2): 200-206. |
朱念德, 刘蔚秋, 伍建军, 等. 1999. 影响南方红豆杉种子萌发因素的研究[J]. 中山大学学报, 38(2): 75-79. |
朱琼琼, 鲁长虎. 2007. 食果鸟类在红豆杉天然种群形成中的作用[J]. 生态学杂志, 26(8): 1238-1243. |
Chambers J C, MacMahon J A. 1994. A day in the life of a seed: movements and fates of seeds and their implications for natural and managed systems[J]. Ann Rev Ecol Syst, 25: 263-292. DOI:10.1146/annurev.es.25.110194.001403 |
DiFazio S P, Vanca N C. 1996. Variation in sex expression of Taxus brevifolia in western Oregon[J]. Canadian Journal of Botany, 74: 1943-1946. DOI:10.1139/b96-232 |
Forget P M, Lambert J, Hulme P E, et al. 2004. Seed fates: seed predation, seed dispersal and seedling establishment[J]. CABI, Publishing, Wallingford, 432. |
García D, Zamora R, Hódar J A, et al. 2000. Yew (Taxus baccata L.) regeneration is facilitated by fleshy-fruited shrubs in Mediterranean environments[J]. Biological Conservation, 95: 31-38. DOI:10.1016/S0006-3207(00)00016-1 |
Giertych P. 2000. Factors determinging natural regeneration of yew (Taxus baccata L.) in the Kórnik Arboretum[J]. Dendrobiology, 45: 31-40. |
Hulme P H. 1996. Natural regeneration of yew (Taxus baccata L.): microsite, seed or herbivore limitation?[J]. Journal of Ecology, 84: 853-861. DOI:10.2307/2960557 |
Johnson R A, Willson M F, Thompson J N, et al. 1985. Nutritional values of wild fruits and consumption by migrant frugivorous birds[J]. Ecology, 66: 819-827. DOI:10.2307/1940543 |
Levey D, Silva W R, Galetti M. 2002. Seed dispersal and frugivory: ecology, evolution and conservation[J]. CABI, Publishing, Wallingford, 511. |
Murray D R. 1986. Seed Dispersal[M]. Sydney: Academic Press.
|
Nathan R, Muller-Landau H C. 2000. Spatial patterns of seed dispersal their determinants and consequences for recruitment[J]. Trends Ecol Evol, 15: 278-285. DOI:10.1016/S0169-5347(00)01874-7 |
Thomas P A, Polwart A. 2003. Biological flora of the british isles Taxus baccata L[J]. Journal of Ecology, 91: 489-524. DOI:10.1046/j.1365-2745.2003.00783.x |