 2010, Vol. 46
2010, Vol. 46文章信息
- 刘爽, 王庆成, 刘亚丽, 田宇明, 孙晶, 徐静
- Liu Shuang, Wang Qingcheng, Liu Yali, Tian Yuming, Sun Jing, Xu Jing
- 土壤pH值对脂松苗木膜脂过氧化及内源保护系统的影响
- Effects of Soil pH Variations on Lipid Peroxidation and Inherent Protection System of Red Pine Seedlings
- 林业科学, 2010, 46(2): 152-156.
- Scientia Silvae Sinicae, 2010, 46(2): 152-156.
-
文章历史
- 收稿日期:2009-03-09
-
作者相关文章



脂松(Pinus resinosa)是北美东北部地区最重要的造林树种之一。因材质优良,生长迅速和耐低温、干旱、瘠薄的特点,被广泛应用于用材林、防风固沙林建设。脂松原产区气候特点和我国东北地区相似,均具有无霜期短、冬季寒冷、夏季温暖、降水量低至中等等特点,且海拔高度相近。我国自2002年在东北地区开始脂松的引种栽培工作,2004年在东北林业大学帽儿山实验林场、辽宁省实验林场和吉林省露水河林业局进行引种栽培试验,目前试验林适应良好,有望在东北地区进行扩大栽培。由于土壤pH值可通过直接和(或)间接途径对植物根系生理生化过程影响直接限制其生长、发育,而成为确定引种植物适生范围的重要因素。
异常的土壤pH值的环境胁迫,对植物的生长发育和生理代谢过程产生影响。目前研究表明:土壤pH值对烟草叶片保护酶活性(王思远等,2005)、丙二醛(MDA)含量(崔喜艳等,2001)、喜树(Camptotheca acuminata)苗期叶片脯氨酸含量(冯建灿等,2001)有显著影响。植物受到环境胁迫,产生生理反应,植物对胁迫的反应强度决定了植物的适应性(武维华,2003)。
关于适宜脂松生长的土壤pH值并不明确。Alban(1974)认为脂松的适生土壤pH值是4.5~6.0,在表土碱性地区不能生长;Hauge等(1991)则认为脂松在土壤pH值5.0~6.5的范围内生长良好;Rawinski等(1992)等认为最适宜脂松苗木生长的土壤pH为5.1~5.5,且在这个范围内土壤pH值和脂松苗木的高生长之间存在明显的负相关;美国农业部林务署森林清查与分析计划书体系的数据显示脂松能够生长的pH值范围为2.8~7.9。以上数据表明:脂松可以生长的土壤pH值最大范围是2.8~7.9,土壤pH4.5~6.5是较适宜的范围,但是没有明确脂松生长的最适宜pH值,也没有表明2.8~4.5和6.5~7.9这2个pH区间苗木的生长受到影响的程度,并且这些数据都来自国外,我们更加需要了解在我国东北地区的特定条件下,脂松苗木的最适土壤pH值范围。
目前,脂松引种栽培区土壤pH范围在5.1~6.5之间,而黑龙江省土壤pH值范围为5.1~8.5(何万云等,1992)。为实现脂松在东北地区的扩大栽培,必须考察其对土壤环境的适应性。本文以2年生脂松苗木为研究对象,采用室内盆栽的方法,研究不同土壤pH值对脂松苗木膜脂过氧化、质膜透性和渗透调节等生理特性的影响,据此对适宜脂松生长的土壤pH值范围进行评价,为其在东北地区的推广栽培提供理论依据。
1 材料与方法 1.1 试验材料土壤取自东北林业大学帽儿山实验林场老山人工林试验站。土壤类型为暗棕壤,在腐殖质层取土。腐殖质土过筛,去除石砾及植物残体,将其与河沙(4:1)的充分混合为栽培基质。栽培基质的碱解氮含量为555.7 mg·kg-1,有效磷含量为37.3 mg·kg-1,速效钾含量为113.1 mg·kg-1,pH值约为5.5。栽培容器为塑料盆(上口径26 cm,下口径16 cm,高13 cm),每盆装入5.00 kg基质。
栽培苗木来自东北林业大学帽儿山实验林场培育的2年生脂松实生苗,共计60盆,每盆栽4株。试验在塑料大棚内进行,每天浇水,保持土壤含水量在田间持水量的60%左右,缓苗15天后开始处理。
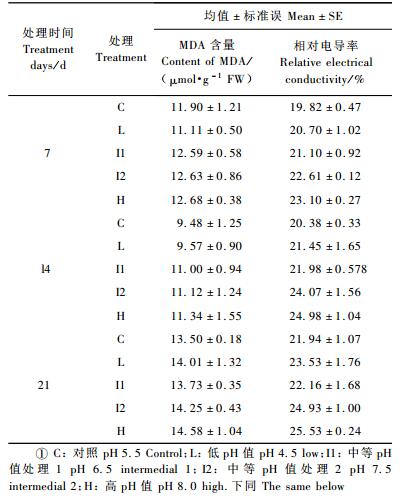
1.2 试验处理根据脂松产区的数据和黑龙江省土壤的pH值情况,设置基质pH值。L组(低pH值): 4.5;C组(对照): 5.5;I1组(中等pH值处理1): 6.5;I2组(中等pH值处理2): 7.5;H组(高pH值): 8.0,并控制各组pH变动在±0.2以内。基质中每天浇不同浓度的NaOH或H2SO4溶液,对照浇自来水(pH 7.1)。处理开始后每天测定1次基质pH值,稳定后(稳定即指连续3天测定的土壤pH值都在设置的范围内)每隔2天测定1次基质pH值,稳定后即可正常浇水,若pH值不在设置范围内时,及时用NaOH或H2SO4溶液调节。基质pH值稳定1周后开始取样,共取样3次,每处理3个重复,取样间隔期为1周。L组中每盆累计浇1 mmol·L-1的H2SO4 10.3 L; I1, I2, H组中每盆分别累计浇1, 2, 3 mmol·L-1的NaOH 13.2, 13.8和10.9 L。
1.3 试验方法5种处理分别随机取样,摘取足量针叶,放入封口袋内,再装入冰盒中,带回实验室。测定针叶的相对电导率,超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)活性和MDA、脯氨酸、可溶性糖含量。
MDA含量采用硫代巴比妥酸(TBA)法测定(李合生等,2000);SOD活性采用氯化硝基四氮唑蓝(NBT)光化还原法测定;CAT活性采用过氧化氢(H2O2)还原法测定;POD活性采用愈创木酚氧化法测定(陈建勋等,2005);类胡萝卜素含量测定: 95%乙醇浸提,浸提液在波长665, 649和470 nm处比色测定吸光度,用Lichtenthaler公式计算类胡萝卜素含量(武维华,2003);相对电导率采用电导仪法测定;脯氨酸含量采用酸性茚三酮溶液显色法测定(汤章诚等,1999);可溶性糖含量采用硫酸-蒽酮比色法测定(王晶英等,2003)。
1.4 数据处理采用SPSS 11.5统计软件处理数据,采用方差分析和最小显著差异法(LSD)比较不同处理间的差异。
2 结果与分析 2.1 基质pH值对脂松苗木针叶丙二醛含量、质膜透性的影响脂松苗木针叶丙二醛含量受时间和基质pH值的影响,且均达到显著水平(P < 0.05),时间和处理的交互作用对其有影响,但未达到显著水平(P>0.05),(表 1)。3次取样,均为对照的MDA含量最低,I2和H组则明显偏高(表 2)。处理7天时,I2,H组MDA含量分别比对照高出6.2%和6.6%;14天时,分别高出17.2%和19.6%;21天时,分别高出5.9%和8.1%(表 2)。
|
|
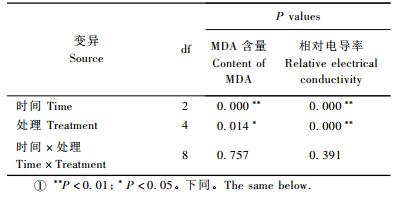
不同时间,不同处理脂松针叶相对电导率差异显著(P < 0.05),时间和处理的交互作用对其也有显著影响(P < 0.05),(表 1)。3次取样,均为对照的相对电导率最低,I2和H组则明显偏高(表 2)。处理7天时,I2,H组相对电导率分别比对照高出14.1%和16.5%;14天时,分别高出18.1%和22.6%;21天时,分别高出13.6%和16.3%(表 2)。
|
|
不同时间,不同处理脂松针叶类胡萝卜素含量、SOD活性、CAT活性差异显著(P < 0.05),时间和处理的交互作用对其也有显著影响(P < 0.05)(表 3)。处理间类胡萝卜素含量、SOD活性、CAT活性的变化趋势均为C> L> I1>I2> H(表 4)。对照的类胡萝卜素含量比L, I1, I2, H分别高出8.41%,24.73%,28.89%和54.67%;对照的SOD活性比L, I1, I2, H分别高出4.78%,7.83%,41.73%和51.19%;对照的类胡萝卜素含量比L, I1, I2, H分别高出4.84%,19.14%,29.56%和40.59%(表 4)。
|
|

|
|
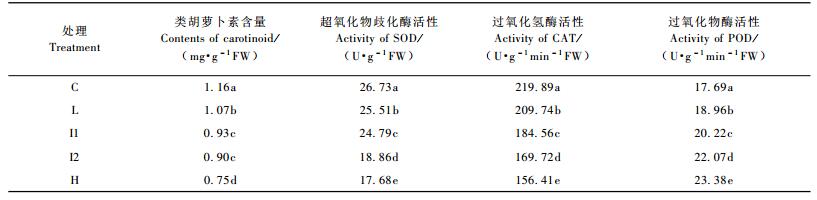
不同时间,不同处理脂松针叶POD活性差异显著(P < 0.05),时间和处理的交互作用对其有影响,但未达到显著水平(P>0.05),(表 3)。与SOD和CAT不同,POD活性H组最高,对照最低,L, I1, I2, H分别比对照高出7.18%,14.30%,24.76%和32.17%(表 4)。
2.3 基质pH值对脂松苗木针叶脯氨酸和可溶性糖含量的影响不同时间,不同处理脂松针叶脯氨酸含量差异显著(P < 0.05),时间和处理的交互作用对其也有显著影响(P < 0.05)(表 5)。对照脯氨酸含量最低,L, I1, I2, H依次升高,分别比对照高出7.03%,19.79%,43.72%和57.00%(表 6)。
|
|
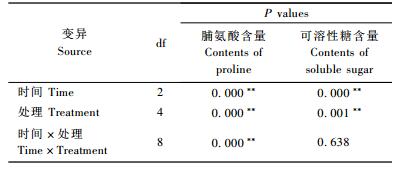
脂松苗木针叶可溶性糖含量受时间和基质pH值的影响,均达到显著水平(P < 0.05),时间和处理的交互作用对其有影响,但未达到显著水平(P>0.05),(表 5)。对照可溶性糖含量最低,L, I1, I2, H依次升高,分别比对照高出5.45%,6.97%,11.76%和15.90%(表 6)。
|
|

逆境导致植物体内氧代谢失调,自由基积累(陈少裕,1991),大量的自由基以极其强烈的氧化作用诱发细胞膜中的不饱和脂肪酸,发生过氧化作用,产物中以MDA含量的增高最为显著(刘丽华等,2007)。本研究中,土壤pH 5.5的处理,MDA含量最低,pH 7.5和pH 8.0的处理含量最高,在处理14天时,pH8.0的处理MDA含量较pH5.5的处理高出19.6%是最大差异(表 2),表明pH 5.5的处理条件下,脂松苗木针叶膜脂过氧化程度最低,土壤pH值大于7.5时,膜脂过氧化程度偏高。崔喜艳等(2001)的研究表明土壤pH值对烤烟叶片内丙二醛含量最大影响可产生37.2%的差距,高于本试验。
细胞膜透性变化是反映细胞膜破坏程度的重要指标(Walter,1997)。电导率的增加说明细胞膜受到潜在的损伤,同时需要指出,20%~25%的电导率(表 2)离质膜的不可逆伤害程度(50%左右)尚远,不足以直接导致细胞和针叶的死亡(杜英君等,2000)。本研究中pH 5.5的处理相对电导率最低(表 2),表明在土壤pH 5.5条件下,细胞膜损伤程度最轻。崔喜艳等(2001)的研究表明:不同pH值处理间烟草叶片的相对电导率差距最大达39.6%,响应较脂松苗木明显。
逆境中,植物组织产生更多的活性氧破坏生物功能分子,SOD, CAT和POD都是对活性氧直接起作用的抗氧化酶类(尹永强等,2007)。在细胞中,SOD把超氧自由基转化为H2O2;CAT能清除大量的H2O2,但与其亲和力较小,不能使细胞内H2O2含量降低到无害水平;POD与H2O2亲和力较大,能清除较低含量的H2O2(吴楚等,2002)。本研究中各处理间SOD和CAT活性比较,均为pH 5.5处理的酶活性最高,pH 7.5和pH 8.0 2处理的酶活性最低(表 4)。SOD活性的高低可反映出植物对所处环境的适应性,不适宜的pH值可引起SOD活性降低(冯建灿等,2001)。本研究3次取样中pH 5.5处理的SOD活性比pH 7.5和pH 8.0 2组处理平均高出41.73%和51.19%(表 4),冯建灿等(2001)研究表明不同pH培养液可使喜树幼苗SOD活性产生16.6%的变化,崔喜艳等(2001)研究表明不同pH值土壤对烟草成熟期SOD活性影响高达732%。
与SOD和CAT活性相反,pH 5.5的处理POD活性最低,且与pH 7.5和pH 8.0处理组差异显著(P < 0.05)(表 3,表 4)。除保护作用外,POD还可能参加活性氧的生产、叶绿素的降解,并能引发膜脂过氧化作用,表现为伤害效应(尹永强等,2007),一般认为以此为主(Zhang et al., 1994)。
类胡萝卜素可清除逆境过程中产生的单线态氧,保护植物叶片(Gilmore, 1997; Reddy et al., 2004)。本研究中,pH值5.5的处理,类胡萝卜素含量最高,表明此处理条件下可能使脂松苗木生长状况较好(表 4)。
膜系统完整性降低,电解质及小分子有机物外渗,膜内外平衡受到破坏,植物细胞积累可溶性物质,进行渗透调节(Reddy et al., 2004; Alonso et al., 2001)。在不同的逆境下,植物代谢反应的总趋势是一致的,即水解作用增强,合成作用减弱,可溶性糖作为渗透调节物质,在植物体内逐步增加(涂三思等,2004)。脯氨酸是1种相溶渗透剂,既作为渗透调节剂维持低水势,又保护生物大分子不受盐离子的毒害,在高浓度下还不影响功能蛋白的结构和活性(许祥明等,2000)。脯氨酸还可以清除细胞的活性氧,减少单线态氧对类囊体膜的光损伤(Reddy et al., 2004; Pinhero et al., 2001)。本研究中可溶性糖和脯氨酸含量都表现pH 5.5处理最低,pH 7.5和pH 8.0处理条件下,脯氨酸含量比对照平均高出43.72%和57.00%(表 6)。冯建灿等(2001)研究指出不同pH培养液可使喜树幼苗脯氨酸含量产生16.6%的变化,崔喜艳等(2001)研究表明不同pH值土壤可使烟草叶片脯氨酸含量发生63%的变化。
综上所述,脂松苗木针叶在土壤pH5.5的条件下,各项生理指标综合表现最佳,其次是pH4.5和6.5,在土壤pH7.5和8.0的条件下,各项生理指标综合表现最差。因此,在我国东北地区的气候土壤条件下,最适宜脂松生长的土壤pH值为5.5,其次为4.5和6.5,pH值7.5以上则不适宜脂松栽培。
陈建勋, 王晓峰. 2005. 植物生理学实验指导[M]. 广州: 华南理工大学出版社.
|
陈少裕. 1991. 膜脂过氧化对植物细胞的伤害[J]. 植物生理学报, 27(2): 84-90. |
崔喜艳, 陈展宇, 王思远, 等. 2001. 土壤pH值对烤烟叶片内超氧物歧化酶活性及丙二醛含量的影响[J]. 吉林农业大学学报, 23(3): 13. |
杜英君, 靳月华. 2000. 远紫外辐射对紫杉幼苗针叶膜脂过氧化及内源保护系统的影响[J]. 应用生态学报, 11(5): 660-664. |
冯建灿, 邓建钦, 张玉洁, 等. 2001. 培养液pH值对喜树幼苗生长与SOD活性、脯氨酸和叶绿素含量的影响[J]. 经济林研究, 19(3): 8. |
何万云, 张之一, 林伯群. 1992. 黑龙江土壤[M]. 北京: 农业出版社.
|
李合生, 孙群, 赵世杰, 等. 2000. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社.
|
刘丽华, 李保国, 齐国辉, 等. 2007. 雄性不育板栗雄花序败育与几种酶活性及MDA含量的关系[J]. 林业科学, 43(4): 121-123. |
汤章诚, 魏家绵, 陈因, 等. 1999. 现代植物生理学实验指南[M]. 北京: 科学出版社.
|
涂三思, 秦天才. 2004. 高温胁迫对黄姜叶片脯氨酸、可溶性糖和丙二醛含量的影响[J]. 湖北农业科学, (4): 100. |
王晶英, 敖红, 张杰, 等. 2003. 植物生理生化实验技术与原理[M]. 哈尔滨: 东北林业大学出版社.
|
王思远, 崔喜艳, 陈展宇, 等. 2005. 土壤pH值对烤烟叶片光合特性及体内保护酶活性的影响[J]. 华北农学报, 20(6): 11-14. |
吴楚, 王政权. 2002. 冰冻条件下外源SA对水曲柳幼苗叶片内抗氧化酶的影响[J]. 林业科学, 38(5): 54-59. DOI:10.11707/j.1001-7488.20020509 |
武维华. 2003. 植物生理学[M]. 北京: 科学出版社.
|
许祥明, 叶和春, 李国凤. 2000. 脯氨酸代谢与植物抗渗透胁迫的研究进展[J]. 植物学通报, 17(6): 536. |
尹永强, 胡建斌, 邓明军. 2007. 植物叶片抗氧化系统及其对逆境胁迫的响应研究进展[J]. 中国农学通报, (1): 105-106. |
Walter L. 1997. 植物生态生理学. 瞿志席, 译. 北京: 中国农业大学出版社.
|
Alonso R, Elvira S, Castillo F J, et al. 2001. Interactive effects of ozone and drought stress on pigments and activities of antioxidative enzymes in Pinus halpensis[J]. Plant Cell Environ, 24(90): 5-16. |
Gilmore A M. 1997. Mechanistic aspects of xanthophylls cycle dependent photoprotection in higher plant chloroplasts and leaves[J]. Physiology Plant, 99: 197-209. DOI:10.1111/ppl.1997.99.issue-1 |
Hauge C T, McMahon B. 1991. Species for Christmas trees in the Lake States[J]. Forestry Facts, 9: 55. |
Pinhero R G, Rao M V, Palyath G, et al. 2001. Changes in the activities of antioxidant enzymes and their relationship to genetic and paclobutrazol-induced chilling tolerance of maize seedlings[J]. Plant Physiology, 114: 695-704. |
Rawinski J J, Bowles J A, Noste N V. 1992. Soil properties related to coniferous seedling height growth in northern Wisconsin[J]. USDA Forest Service, 114(25): 1-3. |
Reddy A R, Chaitanya K V, Vivekanandan M. 2004. Drought-induced responses of photosynthesis and antioxidant metabolism in highter plants[J]. Journal of Plant Physiology, 161(11): 1189-1202. DOI:10.1016/j.jplph.2004.01.013 |
Zhang J X, Kirkham M B. 1994. Drought-stress-induced changes in activities of superoxide dismutase, catalase, and peroxidase in wheat species[J]. Plant Cell Physiology, 35(5): 785-791. DOI:10.1093/oxfordjournals.pcp.a078658 |