 2010, Vol. 46
2010, Vol. 46文章信息
- 陈顺立, 吴晖, 邓招娣, 魏初奖, 黄振裕, 洪贞
- Chen Shunli, Wu Hui, Deng Zhaodi, Wei Chujiang, Huang Zhengyu, Hong Zhen
- 马尾松不同家系营养物质含量与松突圆蚧抗性的关系
- Relationship between Nutrients Content of Different Families of Pinus massoniana and Its Resistance against Hemiberlesia pitysophila
- 林业科学, 2010, 46(2): 87-94.
- Scientia Silvae Sinicae, 2010, 46(2): 87-94.
-
文章历史
- 收稿日期:2008-07-04
-
作者相关文章


2. 福建省森林病虫害防治检疫总站 福州 350003;
3. 三明市森林病虫害防治检疫站 三明 365000
2. General Station of Forest Pest Control and Quarantine in Fujian Province Fuzhou 350003;
3. Forest Pest Control and Quarantine Station in Sanming City Sanming 365000
植物对植食性昆虫的抗性分为组成型抗性和诱导型抗性。组成抗性是植物的一种固有特性,取决于不同基因型,总是存在于植物中并始终起作用;诱导抗性是在表型的水平上考察植物抗性,指被取食的植物影响植食者行为或降低其嗜好性的反应(Agrawal, 1999)。组成抗性涉及植物形态、营养质量、次生性化合物积累对植食性昆虫的负面影响(Smith,1992),而且组成抗性相对比较固定(尤民生等,2000)。因此,组成抗性可以作为树种(或种质)对害虫抗性的早期鉴定指标,可以加快抗虫树种(或种质)抗性育种的进程。诱导型抗性则是在昆虫取食植物时,植物能够调节其体内化学物质的数量或组成,降低植物的营养水平,以对昆虫的取食产生拮抗(Haukioja et al., 1990)。研究植物的诱导抗性,可在实践上补充和完善害虫综合治理理论(娄永根等,1997),而且对减少化学农药或不使用化学农药防治害虫的研究具有重要意义(李镇宇等,2000)。葛振华等(1992)在分析对日本松干蚧(Matsacoccus matsamurae)抗性不同的马尾松(Pinus massoniana)种源的内含物时发现,总糖、游离氨基酸总量以及异亮氨酸、赖氨酸和丙氨酸含量与虫口密度呈正相关。李镇宇等(2000)在油松(P. tabulaeformis)对赤松毛虫(Dendrolimus spectabilis)的诱导化学防御研究中发现,受害油松新生针叶氨基酸总量及多数游离氨基酸含量下降,水溶性总糖、粗脂肪、生物碱含量及总糖/氨基酸比例上升。戈峰等(2002)报道了油松、马尾松、落叶松(Larix)3种松树在受到松毛虫中等程度的危害后,能迅速产生诱导抗性,昆虫生存所必需的还原糖、氨基酸、脂肪酸、蛋白质、Vc等营养物质减少。马尾松受害后产生的滞后诱导抗虫性也与昆虫所需主要营养物质的减少有关。
松突圆蚧(Hemiberlesia pitysophila)是一种危险性极大的国际检疫害虫, 主要危害松树的针叶、嫩梢和球果,使针叶和嫩梢生长受到抑制。松树受害严重时针叶脱落,新抽的枝条变短变黄,甚至导致全株枯死(陈顺立等,2004)。马尾松是受松突圆蚧危害最重的松属树种(陈泽藩等,1988)。马尾松是广泛分布的乡土树种,具有丰富的种质种源。前期研究表明,不同马尾松家系对松突圆蚧的抗性有显著差异(洪贞等,2006)。研究马尾松对松突圆蚧的组成抗性和诱导抗性,有助于筛选抗性强且性状优良的种质以营造抗虫林分,提高松突圆蚧综合控制的效果;还有助于松突圆蚧综合控制过程中减少化学农药的使用。
1 材料与方法 1.1 试验地概况及供试品种试验地设置于福建福清市瑶峰林场,海拔100~220 m,年均气温19.7 ℃,年均降水量1 327.14 mm,全年平均相对湿度77%。试验林中完全随机区组配置马尾松家系30个,无性系7个(设2个区组),树龄均为5年生,造林密度为2 505株·hm-2。地被物主要有铁芒萁(Dicranopteris linearis)、桃金娘(Rhodomyrtus tomentosa)、菝葜(Smilax sp.)等。试验地设有接虫试验林和未接虫试验林,不同处理的试验林以木荷(Schima superba)间隔。
试验林接虫方法:在2004年松突圆蚧盛发期(5月),挑选带有雌成虫的松针束(带虫量每束7~11头)倒插入试验林植株2年生侧梢中上部的针叶束中,每株东、西、南、北4个方向的枝条各接5束,计20束。接虫试验2年后,各供试家系的受害程度和生长状况差异明显,植株出现被害状,部分家系抽梢不正常。家系469达中度发生程度(有虫针束率为31.25%),其余家系有虫针束率小于30%(洪贞等,2006;陈顺立等,2008)。
根据笔者前期研究结果(洪贞等,2006;陈顺立等,2008),选择对松突圆蚧有不同抗性的10个家系进行营养物质的测定。所选家系及其对松突圆蚧的抗性如下:高抗家系:318,330,386,465;中抗家系:155,586,588;低抗家系:439,458,469。
1.2 样品的采集和处理接虫试验林样株于接虫2年后(2006年)采样, 高抗家系平均每束针叶雌成虫数≤1头;中抗家系平均每束针叶雌成虫数>1头、≤3头;低抗家系平均每束针叶雌成虫数≥4头。接虫试验林和未接虫试验林采样方法相同,均为在松突圆蚧盛发期(5月),每家系随机选择6~8株样树,分别于样树上部的东、南、西、北各方向2年生枝条上采集20束未被害的松针,所采松针带回实验室后通过镜检确认无松突圆蚧为害。在相同的区组中,相同家系相同处理的样树上采得的松针混合为1个样品,共采集20份样品。2个区组即为2次重复。一部分样品保鲜带回实验室测定可溶性蛋白含量,其余样品带回实验室后用清水冲洗干净并晾干,然后以60 ℃干燥48 h,粉碎,过40目筛,装袋后置于干燥器中待测。
1.3 营养物质测定方法可溶性糖和总糖含量采用蒽酮比色法测定(吴坤君等,1992;毛燕等,1998)。可溶性蛋白质含量测定采用考马斯亮蓝G-250染色法(路阳等,1992)。全N含量测定采用凯氏定氮法(中国科学院上海植物生理研究所等,1999)。氨基酸含量测定采用离子交换色谱分析法(GB/T 5009.124-2003)。粗脂肪含量测定采用残余法(杨伟,1989)。每个样品的营养物质含量均重复测定2次。
1.4 统计方法采用Mcrosoft Office Excel 2003和DPS软件Ⅴ9.50标准版统计、分析试验数据,多重比较采用Duncan’s新复极差法。各家系针叶营养物质含量的平均值为同处理同家系2个样品平均而得,每个样品的测定值为2次重复测定数据的平均值。
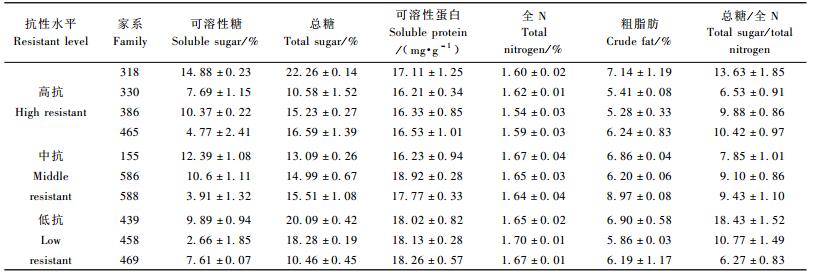
2 结果与分析 2.1 未受害植株营养物质含量各马尾松家系未受害植株针叶的营养物质含量测定结果见表 1、表 2。通过比较3类抗性水平的10个家系的营养物质含量可知,测定的各项营养物质含量中,不同抗性的3类家系之间存在显著差异的只有全N含量(P=0.009 2),高抗家系全N含量平均为1.588%,中抗家系平均为1.653%,低抗家系则为1.673%,新复极差法多重比较结果显示,高抗家系全N含量低于低抗家系,而中抗家系和低抗家系之间,以及中抗家系与高抗家系之间无显著差异(α=0.01)。马尾松各家系总糖含量在10.46%~22.26%之间,高抗家系总糖含量平均值为16.17%,中抗家系平均值为14.53%,低抗家系平均值为16.27%,方差分析显示3类家系总糖含量没有显著差异(P=0.849 4)。家系之间可溶性糖含量差异较大,家系458只含有2.66%的可溶性糖,而家系318则有14.88%,但是3类家系之间可溶性糖含量也没有显著差异(P=0.692 1)。3类不同抗性的家系的粗脂肪和可溶性蛋白含量同样不存在显著差异(P值分别为0.264 1和0.070 6)。随着家系抗性降低,可溶性蛋白含量有增大的趋势。
|
|
|
|
植物的碳氮比(总糖/全N)是衡量食物的营养比例是否适合植食性昆虫的重要指标,在不同抗性的3类家系之间,总糖/全N不存在显著差异(P=0.641 1)。
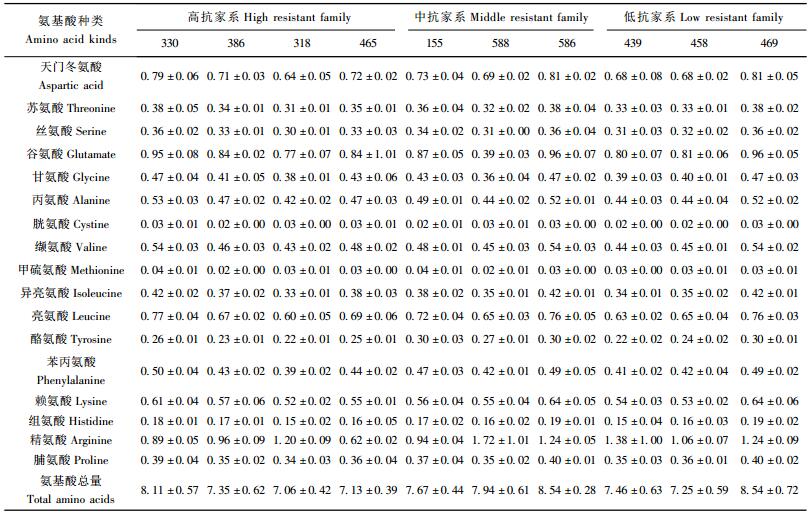
马尾松各家系松针中均测出17种游离氨基酸,其中包括9种昆虫的必需氨基酸。各家系针叶中各种游离氨基酸含量差异很大,相同抗性表现的家系之间各种游离氨基酸含量都大不相同,在3个抗性层次的家系之间各种游离氨基酸含量也不存在显著差异(表 3)。虽然各种游离氨基酸含量在家系之间相差很大,但游离氨基酸总量却相近,在7.06%~8.54%之间。在3个抗性层次的家系之间游离氨基酸总量差异不显著(P=0.370 4)。
|
|

测定结果见表 4、表 5。马尾松植株受松突圆蚧危害后,不同抗性水平家系的可溶性蛋白和全N含量有极显著差异(P值分别为0.000 4和0.008 4),其中高抗家系可溶性蛋白含量(13.335 mg·g-1)极显著低于中抗家系(15.987 mg·g-1)和低抗家系(17.273 mg·g-1)(α=0.01),而中抗家系和低抗家系的可溶性蛋白含量无显著差异。全N含量与家系的抗性之间的关系则略有不同,高抗家系的全N含量极显著低于低抗家系,高抗家系和中抗家系之间无显著差异,低抗家系与中抗家系之间亦无显著差异(α=0.01)。
|
|

|
|
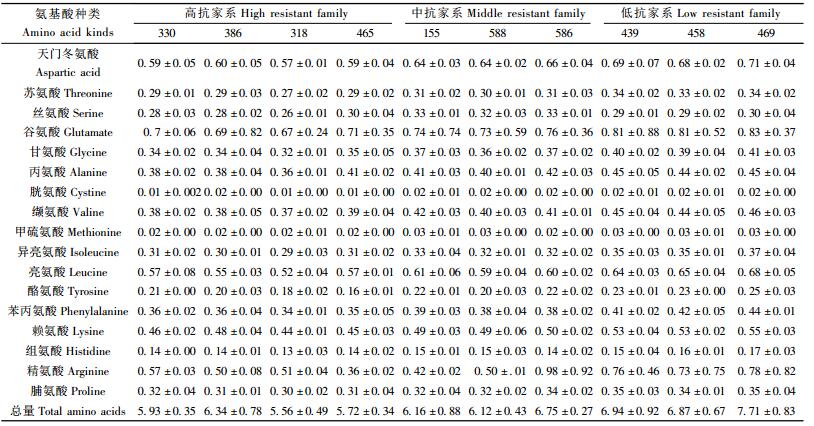
方差分析结果显示,家系对松突圆蚧的抗性不同,其受害植株针叶可溶性糖、总糖、粗脂肪含量以及总糖/全N均无显著差异。
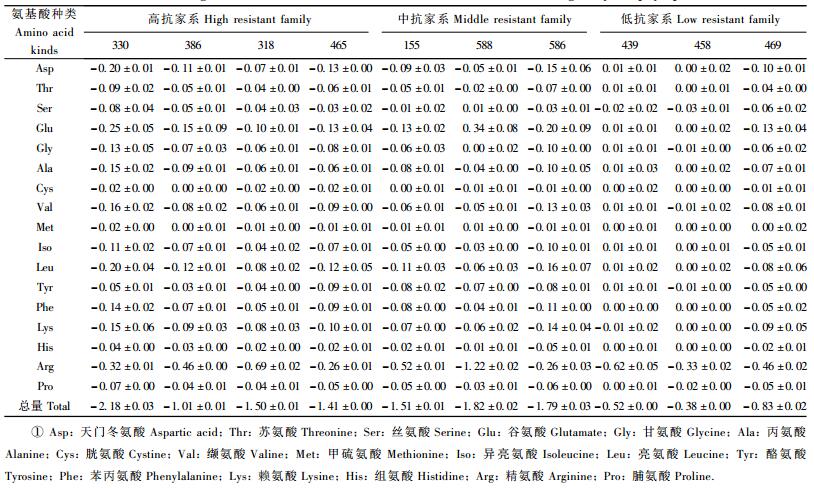
马尾松各家系受松突圆蚧危害后,其针叶中氨基酸种类没有增减,但含量却因家系的抗性而发生了变化,表现为除精氨酸外,其余游离氨基酸含量及总量在不同抗性的3类家系之间存在显著或极显著的差异。除精氨酸外,高抗家系受害植株中松针的16种游离氨基酸含量及总量均低于低抗家系,其方差分析和新复极差法多重比较结果见表 6。
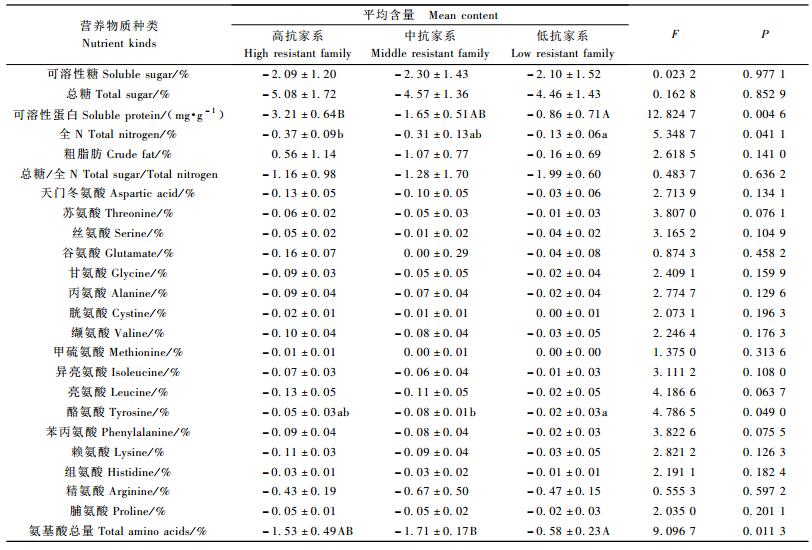
|
|

受松突圆蚧危害后,马尾松各家系的可溶性糖、总糖、可溶性蛋白和全N的含量均有不同程度的降低(表 7)。因试验林中危害程度大部分较轻,故马尾松受到松突圆蚧危害后,针叶营养物质含量下降,应看作是马尾松对松突圆蚧危害产生的诱导反应。在试验中,各个抗性层次家系的植株均发生了这种诱导反应。不同抗性层次家系的可溶性糖、总糖含量的降低量不存在显著差异(P=0.977 1和0.852 9),因此这个糖类物质的诱导反应与家系抗性的高低无关。针叶可溶性蛋白和全N含量也会因松突圆蚧的危害而降低,而且不同抗性层次家系的降低量存在(极)显著差异(P=0.004 6和0.041 1),其中高抗家系可溶性蛋白含量的降低量(3.210 mg·g-1)显著大于低抗家系(0.863 mg·g-1)(α=0.01);高抗家系全N含量降低值(0.370%)显著大于低抗家系(降低值为0.133%,α=0.05)。这说明可溶性蛋白和全N含量的诱导反应在不同抗性层次的家系中存在显著差异,高抗家系反应更强烈。
|
|
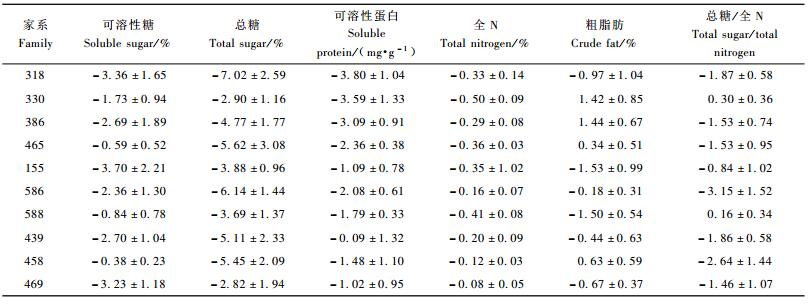
受松突圆蚧危害后,各个家系针叶中游离氨基酸总量都有不同程度的降低,降低量与家系的抗性有关,高抗和中抗家系游离氨基酸总量降低的多(降低量分别为1.525%和1.707%),而低抗家系降低显著较少(降低量为0.577%,P=0.011 3)。
可见,马尾松植株受松突圆蚧危害后会发生营养物质含量降低的诱导反应,其中糖类物质含量的降低与家系对松突圆蚧的抗性高低无关,而可溶性蛋白质、游离氨基酸和全氮等含N物质含量的降低与家系的抗性有关,高抗家系含N物质含量降低较低抗家系多。
粗脂肪含量和总糖/全N则有增有减,方差分析表明,不同抗性层次家系间粗脂肪含量和总糖/全N的变化量不存在显著差异(P=0.141 0和0.636 2)。
大多数游离氨基酸的含量在植株受到松突圆蚧的危害后或降低,或无明显变化(表 8)。除酪氨酸外的其余16种游离氨基酸含量因植株受害导致的变化在3个不同抗性层次的家系之间均不存在显著差异。在3个抗性层次的家系植株中,酪氨酸含量的降低值存在显著差异,中抗家系受害后,酪氨酸含量降得比低抗家系多(P=0.049 0)(表 9)。
|
|
|
|
陈泽藩等(1988)研究表明,马尾松是15种松属树种中对松突圆蚧抗性最弱的一种,相同条件下,其受松突圆蚧危害最重。陈顺立等(2008)、洪贞等(2006)研究表明,马尾松虽然易受松突圆蚧的危害,但也存在着对松突圆蚧有较强抗性的家系。本文从不同抗性的家系受松突圆蚧危害前后营养物质的含量及变化情况,初步探讨了马尾松家系对松突圆蚧的抗性机制。
研究结果表明,家系对松突圆蚧的抗性与针叶中营养物质含量有关。高抗家系针叶中全N含量低构成了马尾松组成抗性的一部分,含N化合物含量高的家系对松突圆蚧的抗性较低。同时,笔者认为松突圆蚧危害诱导马尾松营养物质含量发生变化,则是马尾松诱导抗性的组成部分。松突圆蚧诱导产生的马尾松营养物质含量变化有:1)针叶中糖类物质含量降低,这个诱导反应在对松突圆蚧有不同抗性的马尾松家系中普遍发生,其反应的强度及受害植株中糖类物质含量多少与家系抗性的高低无关;2)针叶中全N含量、可溶性蛋白含量和游离氨基酸总量等含N化合物含量也有不同程度的降低。这些含N化合物含量降低的程度与家系对松突圆蚧抗性的高低有关,高抗家系降低量大于低抗家系;3)高抗家系受害植株针叶中16种游离氨基酸含量低于低抗家系。
营养物质可能在下述情况下引起植物对植食性昆虫的抗生作用:1)缺乏某些营养物质,如维生素或主要氨基酸;2)某些营养物质,特别是氨基酸或特定的固醇含量不足;3)有效营养物质不平衡(Painter,1969)。松突圆蚧连年在马尾松各个家系上危害,表明马尾松各个家系的营养物质种类和含量均能够满足松突圆蚧生长发育的需求,所以马尾松对松突圆蚧的抗性不是因为营养物质的缺乏。其次,研究表明,针叶中C/N在不同抗性层次的家系间没有显著差异,说明马尾松对松突圆蚧的抗性不是通过改变营养物质比例而实现的。因此,马尾松对松突圆蚧的抗生作用是针叶中某些营养物质含量的降低,从而抑制了松突圆蚧的正常生长发育。马尾松受松突圆蚧危害后,可溶性糖、总糖、可溶性蛋白、全N、总游离氨基酸等营养物质含量降低,说明这些营养物质的不足或下降可引起马尾松对松突圆蚧的抗生作用。
糖类物质、可溶性蛋白、氨基酸对昆虫的营养作用已较明确(彩万志,2001)。N是植物体内许多重要有机化合物的组分,其中蛋白态N通常占80%~85%,核酸态N约占10%,此外,N还是叶绿素、一些维生素和许多酶的组分(陆景陵,1994)。有关哪些含N化合物在马尾松对松突圆蚧抗性中起作用及其作用的机制还有待进一步研究。
GB/T5009. 124-2003. 食品中氨基酸的测定方法. 中华人民共和国国家标准.
|
Smith C M. 1992. 植物抗虫性的研究与应用. 冯明光, 译. 北京: 中国农业科技出版社, 292.
|
彩万志. 2001. 普通昆虫学[M]. 北京: 中国农业大学出版社: 118-120.
|
陈顺立, 林庆源, 黄金聪. 2004. 南方主要树种害虫综合管理[M]. 厦门: 厦门大学出版社: 113-121.
|
陈顺立, 吴晖, 洪贞, 等. 2008. 马尾松家系受松突圆蚧危害后生长状况研究[J]. 福建林学院学报, 28(2): 97-100. |
陈泽藩, 杨肇兴, 徐家雄, 等. 1988. 十五种松树对松突圆蚧抗性的初步研究[J]. 森林病虫通讯, (2): 1-2. |
戈峰, 李镇宇, 谢映平, 等. 2002. 我国主要松树诱导抗虫性的一些规律比较[J]. 北京林业大学学报, 24(3): 61-65. |
葛振华, 陆琴华. 1992. 马尾松抗蚧种源的皮型和内含物分析[J]. 林业科学研究, 5(4): 483-486. |
洪贞, 陈顺立, 吴晖, 等. 2006. 松突圆蚧对不同马尾松家系危害的初步研究[J]. 华东昆虫学报, 15(3): 201-205. |
李镇宇, 王燕, 陈华盛, 等. 2000. 油松对赤松毛虫的诱导化学防御及滞后诱导抗性[J]. 林业科学, 36(1): 66-70. |
娄永根, 程家安. 1997. 植物的诱导抗虫性[J]. 昆虫学报, 40(3): 321-327. |
陆景陵. 1994. 植物营养学[M]. 北京: 北京农业大学出版社.
|
路阳, 王培之, 丁丽俐. 1992. 用考马斯亮蓝G-250迅速灵敏地测定蛋白质浓度[J]. 生物学杂志, 45(1): 24-26. |
毛燕, 王崇利. 1998. 毛竹等九种竹叶中蛋白质和总糖含量的测定[J]. 竹子研究汇刊, 17(2): 18-20. |
吴坤君, 李明辉. 1992. 棉铃虫营养生态学的研究:食物中糖含量的影响[J]. 昆虫学报, 35(1): 47-52. |
杨伟. 1989. 残余法测定香椿籽中粗脂肪含量的研究[J]. 山东农业大学学报:自然科学版, (4): 75-78. |
尤民生, 侯有明, 魏辉, 等. 2000. 植物诱导防御的空间和时间效应[J]. 武夷科学, 16: 195-201. |
中国科学院上海植物生理研究所, 上海市植物生理学会. 1999. 现代植物生理学实验指南[M]. 北京: 科学出版社: 133-134.
|
Agrawal A A.1999.Induced plant defense:evolution of induction and adaptive phenotypic plasticity//Agrawal A, Tuzun S, Bent E, et al.Inducible plant defenses against pathogens and herbivores:Biochemistry, ecology, and agriculture.St.Paul, Minnesota:American Phytopatholosical Society Press, 251-268.
|
Haukioja E, Neuvomen S. 1990. Theories of plant chemical defense in trees[J]. Ann Rev Entomol, 36: 25-42. |
Painter R H. 1969. Plant and animal resistance to insect[J]. Principles of Plant and Animal Pest Control, (3): 64-69. |