 2010, Vol. 46
2010, Vol. 46文章信息
- 赵志红, 吴红芝
- Zhao Zhihong, Wu Hongzhi
- 月季切花水分胁迫耐性差异与脯氨酸的关联性
- Relationship between Tolerance Difference to Water Stress and Proline Content in Cut Rose Cultivars
- 林业科学, 2010, 46(2): 74-79.
- Scientia Silvae Sinicae, 2010, 46(2): 74-79.
-
文章历史
- 收稿日期:2009-01-12
-
作者相关文章



月季(Rosa hybrida)切花作为世界四大切花之一, 在国内外花卉市场占有非常重要的地位。随着月季切花生产向自然气候条件优越、劳动力成本低地区转移的发展趋势,月季切花远距离运输变得越来越普遍。目前国内外月季切花的远距离运输多采用没有任何水分补充措施的干运方式,水分胁迫引起的僵蕾、僵花、弯颈、萎蔫等质量损失成为限制月季切花远距离运输品质的主要因子之一(赵喜亭等,2008)。此外,月季切花从采收、预冷、包装以及贮藏等各个采后环节中,都有可能遭到不同程度的失水胁迫(Mayak et al., 1971; 1985),制约了月季切花生产的发展(Zieslin,1989;Urban et al., 2002)。目前有关月季切花水分胁迫耐性机理的研究,主要集中在失水胁迫与抗氧化酶系统和非酶系统以及与内肽酶的变化之间的关联等方面(张常青等,2002;丛日晨等,2003a; 2003b;刘晓辉等,2005;赵喜亭等,2008),有关细胞渗透调节物质变化与水分胁迫耐性方面的研究尚未见报道。本试验通过对目前市场流行的12个月季切花品种水分胁迫耐性的比较,筛选出对水分胁迫耐性高和敏感的品种,并进一步研究耐性和敏感品种之间细胞渗透调节物质脯氨酸含量的差异, 探索月季切花水分胁迫耐性机理,为月季切花耐性品种的筛选及远途运输保鲜技术的制定提供参考。
1 材料与方法试验于2007年9月—2008年9月在云南农业大学鲜切花采后生理实验室进行。
1.1 试验材料供试的12个月季切花品种,由昆明杨月季园艺有限公司提供(表 1)。采收时间为上午9:00,采收长度50 cm,开花指数为2级(马男等,2005)。
|
|
所有月季切花采收后插入自来水中运回实验室,按花枝长45 cm,留取3~4片叶, 下端浸入清水中45°斜面进行修剪。品种筛选共设4组水分胁迫处理,即胁迫24 h(a),36 h(b),48 h(c)和对照(ck);筛选出来的水分胁迫耐性品种和敏感品种进行7组水分胁迫处理,即胁迫6,12,18,24,30,36 h和对照,每项测定每个处理取花材10支,重复3次, 处理后复水瓶插观测,对照花材一直插养于水中作同样观测。各种水分胁迫处理花材在室温22.5~27 ℃(平均温度为24.7 ℃),相对湿度在44%~64%(平均湿度为56.7%)的条件下进行。胁迫后复水瓶插,瓶插时白天日光灯补光(光照强度为25.7 μE·m-2s-1)。
1.2.2 水分胁迫评估指标的测定1) 瓶插寿命与瓶插寿命缩短百分率 瓶插寿命指从瓶插之日起到花瓣出现萎蔫、弯头或蓝变前1天的天数。瓶插寿命缩短百分率指胁迫后切花的瓶插寿命与对照的差值占对照切花瓶插寿命的百分率。2)花朵弯颈率与复水率 花朵弯头指花梗部弯软使花头弯下,是切花瓶插寿命结束标志。胁迫后弯颈率指胁迫后弯颈的花枝数占总花枝数的百分率。复水率指失水胁迫后在水中吸水恢复新鲜的花枝数占总花枝数的百分率。3)鲜样质量损失率 鲜质量损失率(%)=(胁迫前鲜质量-胁迫后鲜质量)/胁迫前鲜质量×100%。4)花朵直径 用游标卡尺测花朵的最大直径。每个处理测10支,取平均值。5)脯氨酸含量 采用酸性茚三酮显色法测定(叶尚红,2004)。胁迫期间定期取样,每次取6支,单枝重复。所有统计分析用DPS3.01分析。
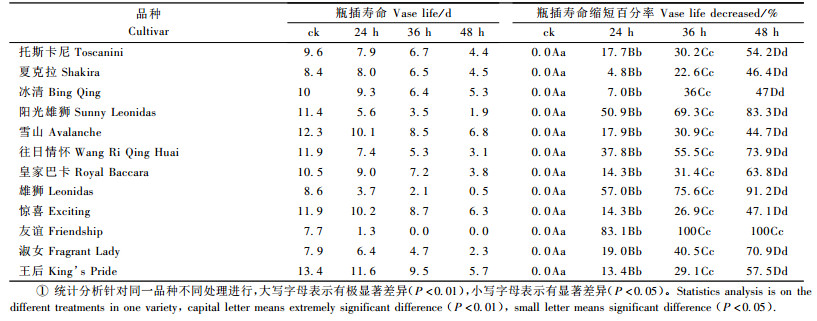
2 结果与分析 2.1 不同品种水分胁迫耐性比较 2.1.1 水分胁迫对瓶插寿命的影响不同品种切花月季在水分胁迫过程中,随着胁迫时间的增加,瓶插寿命呈现缩短趋势,而瓶插寿命缩短百分比呈增加趋势。水分胁迫24 h后,‘阳光雄狮’、‘雄狮’和‘友谊’等3个品种缩短了5.8~6.4天, 缩短百分比为50.9%~83.1%;‘托斯卡尼’、‘雪山’、‘往日情怀’、‘皇家巴卡’、‘惊喜’、‘淑女’和‘王后’等7个品种瓶插寿命缩短了1.8~4.5天, 缩短百分比为13.4 %~37.8%;‘夏克拉’和‘冰清红’等2个品种缩短最少, 为0.4~0.7天, 缩短百分比为4.8%~7.0%。由此可见:水分胁迫对不同切花月季品种瓶插寿命的影响不同, 有的品种瓶插寿命显著缩短; 有的则变化很小。品种‘友谊’的瓶插寿命缩短百分率最高,达83.1%,显著高于变化仅4.8%的‘夏克拉’品种,说明不同的切花月季品种间存在着对水分胁迫耐性的差异。对‘夏克拉’水分胁迫时间由24 h增加到48 h, 瓶插寿命缩短百分率由4.8%增加到46.4%;其他品种也具有相同的趋势(表 1)。但在相同水分胁迫处理时间条件下, ‘友谊’的瓶插寿命缩短百分率大于‘夏克拉’,这说明‘友谊’是对水分胁迫敏感的品种,‘夏克拉’更加耐失水胁迫。
2.1.2 水分胁迫对不同月季切花弯颈率和复水率的影响水分胁迫24 h时,‘夏克拉’和‘皇家巴卡’弯颈率已达到100%,‘托斯卡尼’、‘冰清’、‘雪山’、‘友谊’和‘王后’未出现弯颈现象,‘阳光雄狮’、‘往日情怀’、‘雄狮’、‘惊喜’和‘淑女’5个品种出现不同程度的弯颈现象;胁迫36 h时,‘往日情怀’、‘惊喜’、‘淑女’和‘王后’4个品种弯颈率也达到100%,其他品种的弯颈率较24 h都有所增加;胁迫增加到48 h时,‘雪山’的弯颈率仍为10%,‘友谊’的弯颈率为30%,其余10个品种的的弯颈率都达到了70%以上。说明‘夏克拉’和‘皇家巴卡’的颈部较不耐失水,‘雪山’和‘友谊’的颈部耐失水程度较高(表 2)。
|
|
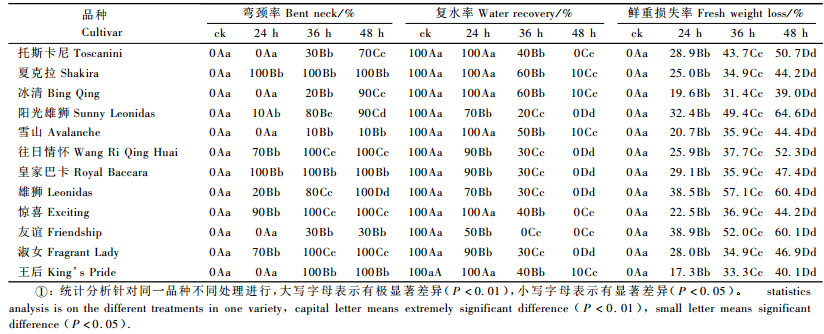
月季切花水分胁迫复水后, 品种之间水分恢复情况差异很大。‘夏克拉’水分胁迫时虽然弯颈现象严重,但复水瓶插后,它的复水能力极强。胁迫24 h时,花枝100%弯颈,复水率达100%;胁迫增加到36 h时,复水率为60%,与‘冰清’一样为最高复水率,而‘友谊’复水率已降为0%;胁迫48 h时,大多数品种的复水率已为0,而‘夏克拉’仍有10%的复水率。由弯颈率和复水率2个指标可知‘夏克拉’的耐失水能力高于其他品种(表 2)。
2.1.3 水分胁迫对不同月季切花鲜重损失率的影响如表 2所示,水分胁迫处理后, 随着水分胁迫加深,鲜重损失率逐渐增加。胁迫24 h后,鲜重损失率最小的是‘王后’,为17.3%;最大的是‘友谊’,为38.5%;‘夏克拉’为25.0%;胁迫36 h后,鲜重损失率最小的是‘冰清’,为31.4%;最大的是‘雄狮’,为57.1%;‘友谊’为52.0%, ‘夏克拉’为34.9%;胁迫48 h后,切花鲜重损失率较胁迫36 h有增加,但变化不大,可能是切花已经在胁迫36 h后,水分已经损失较多,到胁迫48 h时,切花已没有多少水分,鲜重损失率变化较小。表明在相同失水胁迫条件下, ‘王后’、‘夏克拉’、‘冰清’等品种水分亏缺程度较轻, 比‘友谊’、‘雄狮’等品种更具有强的失水胁迫耐性。
2.1.4 水分胁迫对不同月季切花花朵直径的影响如图 1所示,月季切花经胁迫处理后,促进了切花开放,瓶插第1天花径比对照高。经24 h胁迫处理后,‘阳光雄狮’、‘雄狮’和‘友谊’3个品种瓶插天数较之对照明显缩短,花朵直径与对照差异明显,无法开到最大花径,一些样品出现僵蕾现象,而‘夏克拉’等其余9个品种基本可以达到完全开放;36 h后,‘托斯卡尼’、‘阳光雄狮’、‘雄狮’和‘友谊’4个品种呈现僵花现象;48 h后, ‘友谊’和‘雄狮’完全不能复水,瓶插天数缩短明显,花朵不能开放,出现了明显的僵花现象, 失水胁迫已对花枝造成不可逆的伤害。‘夏克拉’、‘冰清’、‘惊喜’、‘淑女’和‘王后’5个品种的对照与3组水分胁迫处理花径变化趋势比较相似,先增加,当花朵直径增加到一定大小时,保持花朵直径大小几乎不改变的状态,直到花朵萎蔫。因此,‘夏克拉’等品种耐水分胁迫能力强于‘友谊’等品种。

|
图 1 水分胁迫对不同月季切花品种花朵直径的影响 Figure 1 Effects of water stress on flowers diameter of different cut rose cultivars |
通过以上的不同品种水分胁迫耐性比较可知:品种‘夏克拉’是其中最耐胁迫的品种,‘友谊’是最敏感的品种。
2.2 水分胁迫时‘夏克拉’和‘友谊’切花中脯氨酸含量的变化随着水分胁迫程度的增加,耐性品种和敏感品种花朵、叶片的游离脯氨酸含量总体是呈逐渐递增,但是达到一定水分胁迫程度后又下降的趋势,且均比对照高,但耐性不同的品种游离脯氨酸积累情况差异十分明显(图 2)。敏感品种‘友谊’在水分胁迫12 h后, 花朵、叶片的游离脯氨酸含量迅速上升;水分胁迫18 h时,花朵脯氨酸含量达到最大值(191 μg·g-1);叶片脯氨酸含量在水分胁迫24 h时达到最大值(208.7 μg·g-1),随后迅速下降。耐性品种‘夏克拉’花朵、叶片的游离脯氨酸含量,随水分胁迫程度的加深持续增加,当水分胁迫36 h时,其含量已远远高于‘友谊’,说明耐性品种细胞的渗透调节能力更大,水分胁迫耐性更强。耐性品种‘夏克拉’和敏感品种‘友谊’经过水分胁迫处理后,叶片、花朵中游离脯氨酸的含量均大于对照,说明月季切花在受到水分胁迫时,游离脯氨酸的积累量与其水分胁迫耐性之间存在正相关性。

|
图 2 水分胁迫对月季切花脯氨酸的影响 Figure 2 Effects of water stress on proline accumulation of cut rose cultivars |
6 h胁迫处理的‘夏克拉’和‘友谊’, 花朵、叶片游离脯氨酸的含量均高于对照, 但‘友谊’比‘夏克拉’增加的幅度大,说明敏感品种‘友谊’对水分胁迫非常敏感, 轻度失水即需大量积累脯氨酸以调节水分平衡;而36 h胁迫处理后, ‘夏克拉’花朵、叶片中游离脯氨酸含量比‘友谊’增加的幅度大。总之,在轻度水分胁迫时, 敏感品种积累更多的游离脯氨酸, 耐性品种则反之,积累量较少;深度水分胁迫时,耐性品种持续积累,脯氨酸含量显著高于敏感品种;敏感品种的脯氨酸积累比耐性品种的反应更快,但耐性品种脯氨酸积累反应的时间更长,具有更强的水分胁迫调节能力。
3 结论与讨论 3.1 不同品种水分胁迫耐性比较月季切花品种种类繁多,品种之间水分胁迫耐性存在着较大的差异(唐雪梅等,1997),贮藏和远距离运输宜选用水分胁迫耐性强的品种,敏感品种只能用于就近销售。瓶插寿命是评价切花品质的重要指标(熊运海,2000),依唐雪梅等(1999)方法,将目前市场流行的12个月季切花新品种的耐失水胁迫能力进行分类,‘阳光雄狮’、‘雄狮’和‘友谊’等3个品种为不耐失水胁迫品种;‘托斯卡尼’、‘雪山’、‘往日情怀’、‘皇家巴卡’、‘惊喜’、‘淑女’和‘王后’7个品种为中度耐失水胁迫品种;‘夏克拉’和‘冰清’2个品种,为耐失水胁迫品种。本试验结果与其划分基本一致。
各种切花失水胁迫后能够恢复的极限不同,相同失水胁迫程度对不同切花月季品种的影响不同;不同失水胁迫程度对同一品种花材的开放和衰老影响也不同,表现为失水胁迫程度较轻时能够恢复,程度较重时无法恢复(唐雪梅等,1997)。在月季切花流通中,赵喜亭等(2005)用快速、便捷的花枝复水恢复率作为衡量品种间的失水胁迫耐性的判断指标。‘夏克拉’水分胁迫时虽然弯颈现象严重,但复水瓶插后,它的复水能力很强。依据赵喜亭等(2005)以复水率为失水胁迫耐性指标,进一步说明‘夏克拉’的耐失水能力高于其他品种。
鲜重损失率是表示植物水分亏缺程度的指标(张常青等,2002)。正常瓶插条件下月季切花花枝鲜重先逐渐增加,达到最大值后再逐渐降低。但是当失水量超过吸水量时花朵开放受到影响,失水量超过一定程度时花朵无法正常开放,出现僵花、僵蕾、萎蔫、弯头等现象,严重降低切花采后品质(高俊平等,1995)。本试验表明在相同失水胁迫条件下, ‘夏克拉’、‘冰清’、‘雪山’和‘王后’4个品种水分亏缺程度较轻, 比‘阳光雄狮’、‘雄狮’和‘友谊’3个品种具有更强的失水胁迫耐性。月季切花随着胁迫时间的延长,鲜重损失率逐渐增加,瓶插寿命缩短。
花朵直径由小变大继而变小,反映了花朵由初开、盛开至萎蔫衰老的进程(白双义等,2001)。经水分胁迫处理瓶插后,‘夏克拉’、‘冰清’、‘惊喜’、‘淑女’和‘王后’5个品种在不同水分胁迫时间下的变化趋势比较相似,花朵直径由小变大, 当花径增大到一定程度时, 月季切花达到盛花状态, 随后一直保持开放状态, 其花径不变小, 直至萎蔫;而‘阳光雄狮’、‘雄狮’和‘友谊’等3个品种水分胁迫处理后花径大小变化剧烈, 瓶插寿命明显缩短。可见月季品种对水分胁迫耐性的差异也能反映在花朵直径的变化上, 因此‘夏克拉’等5个品种耐水分胁迫能力强于‘阳光雄狮’等3个品种。
3.2 不同水分胁迫耐性月季切花与脯氨酸的关联性脯氨酸是植物体内的1种重要渗透调节物质,对干旱胁迫反应较敏感。脯氨酸的积累是植物对水分亏缺的生理反应,累积数量的多少可作为衡量作物抗旱力的生理指标。试验结果表明:月季切花在失水不太严重时, 游离脯氨酸积累少的品种耐失水胁迫, 游离脯氨酸积累多的不耐失水胁迫。失水严重时,耐失水胁迫品种脯氨酸积累的时间长,含量继续增多,即敏感品种‘友谊’比耐性品种‘夏克拉’在水分胁迫下的脯氨酸积累反应更快,而耐性品种‘夏克拉’反应的持续时间较长。这与王启明等(2005)用大豆(Glycine max)苗和汤章成等(1986)用高粱(Sorghum)研究的结果是一致的。至于引起这种差异的原因可能是由于失水胁迫耐性不同的月季切花品种存在着对失水胁迫反应敏感程度不同的脯氨酸代谢调节机制。耐性品种‘夏克拉’和敏感品种‘友谊’经过水分胁迫处理后,叶片、花朵中游离脯氨酸的含量均大于对照。因此,在失水胁迫时,月季切花中游离脯氨酸的含量与其失水胁迫耐性存在有一定的正相关性,可以作为筛选水分胁迫耐性品种的生理指标之一。
白双义, 刘青林. 2001. 月季切花不同品种衰老征兆及瓶插寿命的比较[J]. 园艺学报, 28(4): 364-366. |
蔡永平, 聂凡, 张鹤英, 等. 2000. 水杨酸对月季切花的保鲜效果和生理作用[J]. 园艺学报, 27(3): 228-230. |
丛日晨, 赵喜亭, 刘晓辉, 等. 2003a. 月季切花采后花瓣内肽酶活性的变化[J]. 园艺学报, 30(2): 232-235. |
丛日晨, 赵喜亭, 高俊平. 2003b. 失水胁迫对切花月季'贝拉米'内肽酶的影响[J]. 园艺学报, 30(3): 352-354. |
高俊平, 孙自然. 1995. 我国远距离流通中减少损耗的基本途径[J]. 北京农业大学学报(增刊), 21: 84-88. |
刘晓辉, 朱旭晖, 赵喜亭, 等. 2005. 两个切花月季品种花朵开放和衰老对乙烯的反应及其与内肽酶的关联[J]. 中国农业科学, 38(3): 589-595. |
马男, 蔡蕾, 高俊平, 等. 2005. 外源乙烯对月季(Rosa hybrida)切花花朵开放的影响与乙烯生物合成相关基因表达的关联[J]. 中国科学C辑:生命科学, 35(2): 104-114. |
汤章成, 王育启, 吴亚华, 等. 1986. 不同抗旱品种高粱苗中脯氨酸累积的差异[J]. 植物生理学报, 12(2): 154-162. |
唐雪梅. 1997. 月季切花不同品种水分胁迫耐性差异的生理研究. 北京: 中国农业大学硕士学位论文.
|
唐雪梅, 高俊平, 孙自然, 等. 1999. 切花月季品种失水胁迫耐性差异及忍耐极限初探[J]. 园艺学报, 26(1): 41-48. |
王启明, 马原松. 2005. 不同抗旱品种大豆苗中脯氨酸累积的差异[J]. 商丘职业技术学院学报, 2(4): 63-64. |
熊运海. 2000. 干藏过程中月季切花衰老的生理机制初探[J]. 西南师范大学学报:自然科学版, 25(2): 164-168. |
叶尚红. 2004. 植物生理生化试验教程[M]. 昆明: 云南科技出版社.
|
余红英, 邓世媛, 尹国强, 等. 2004. 保水剂对月季切花生理生化特性的影响研究初报[J]. 广东农业科学, 3(3): 28-29. |
张常青, 唐雪梅, 高俊平, 等. 2002. 月季切花'萨蔓莎'和'加布里拉'失水胁迫耐性的差异[J]. 园艺学报, 29(6): 556-560. |
赵喜亭, 丛日晨, 单宁伟, 等. 2005. 切花月季失水胁迫耐性差异与内肽酶活性的关联[J]. 园艺学报, 32(5): 854-860. |
赵喜亭, 丛日晨, 刘晓静, 等. 2008. 失水胁迫对月季花瓣内肽酶活性的诱导及对花朵衰老进程的影响[J]. 中国农业科学, 41(2): 516-524. |
Mayak S, Halevy A H. 1971. Water stress as the cause of failure of flower bud opening in Iris[J]. Journal of American Society Horticultural Science, 96: 482-483. |
Mayak S, Borochov A, Tirosh T. 1985. Transient water stress in carnation flowers: Effect of Amino-oxyacetic acid[J]. Journal of Experimental Botany, 166: 800-806. |
Urban L, Six S, Barthélémy L, et al. 2002. Effect of elevated CO2 on leaf water relations, water balance and senescence of cut roses[J]. Journal of Plant Physiology, 159(7): 717-723. DOI:10.1078/0176-1617-0602 |
Zieslin N, Kohl H C, Kofranek Jr A M, et al. 1978. Changes in the water status of cut roses and its relationship to bent-neck phenomenon[J]. Amer Soc Hort Sci, 103(2): 176-179. |
Zieslin N. 1989. Postharvest control of vase life and senescence of rose flowers[J]. Acta Horticulture, 261: 257-264. |