 2010, Vol. 46
2010, Vol. 46文章信息
- 周曙光, 孔祥生, 张妙霞, 王丽亚, 王福云, 周桂勤
- Zhou Shuguang, Kong Xiangsheng, Zhang Miaoxia, Wang Liya, Wang Fuyun, Zhou Guiqin
- 遮光对牡丹光合及其他生理生化特性的影响
- Effects of Shading on Photosynthesis, and Other Physiological and Biochemical Characteristics in Tree Peony
- 林业科学, 2010, 46(2): 56-60.
- Scientia Silvae Sinicae, 2010, 46(2): 56-60.
-
文章历史
- 收稿日期:2008-10-31
-
作者相关文章



近年来,牡丹(Paeonia suffruticosa)栽培面积不断扩大,牡丹产业在地方经济中的作用日益凸现。众所周知,光是植物光合作用的能量来源,但是,光能过剩也会抑制光合作用,甚至引起光合机构的光氧化破坏(Demmig-Adams et al., 1992)。周治国等(2001)研究报道,遮荫后棉苗茎叶结构以及功能叶片叶绿体的形态和超微结构发生变化,导致棉花(Gossypium)功能叶片叶绿素含量、光化学效率及光合速率的变化。张国斌等(2004)、刘卫琴等(2006)对辣椒(Capsicum)、草莓(Fragaria)进行遮光试验,发现遮光后植物的光合特性、叶绿素荧光特性等都发生一系列变化。牡丹花农的栽培经验表明,适当遮荫有利于牡丹生长发育,进而影响牡丹成花。但是,有关光强对牡丹生长特性的影响,尤其定量研究光强与牡丹光合及其他生理生化特性关系鲜见报道。本研究旨在探明不同光强条件下牡丹光合及其他生理生化特性的变化,探明对牡丹生长发育最有利的光照强度,为牡丹优化栽培提供理论依据。
1 材料与方法 1.1 试验设计试验在洛阳国际牡丹园和河南科技大学农学与生物技术实验室进行。试验材料为洛阳红牡丹品种,选株龄4年、生长良好、植株大小一致的牡丹,按株、行距均为70 cm种植。2007年3月25日搭棚遮光,设4个处理(Ⅰ:不遮光,Ⅱ:遮光30%,Ⅲ:遮光50%,Ⅳ:遮光80%),每处理8株。5月8日(晴)选第3片叶(自上而下)测定光合和荧光参数的日变化,5月9日上午11:00(晴朗)取第3片叶测定叶绿素、MDA、O2·、总酚、类黄酮和谷胱甘肽含量及SOD和POD的活性。
1.2 测定方法 1.2.1 光合参数测定光合参数用英国产CIRAS-1型便携式光合测定系统测定,主要包括光合有效辐射(PAR)、净光合速率(Pn)和细胞间CO2浓度(Ci)等生理生态因子。测定从8:30开始,11:00,13:00,15:00和17:00左右共进行5轮测定,每轮每处理重复测5次,结果为所测数据的平均值。
1.2.2 叶绿素荧光参数测定叶绿素荧光参数用英国Hansatech公司产的便携脉冲调制式荧光仪测定,最大光化学效率(Fv/Fm)和实际光化学效率(ФPSⅡ)直接由仪器测出。测定从7:00开始,8:00,10:00,12:00,14:00,16:00和18:00左右共进行7轮测定,每轮每处理重复测定5次,结果为所测数据的平均值。
1.2.3 酶活性测定SOD活性测定采用NBT光还原法(王宪泽等,2002),POD活性采用愈创木酚法(高俊凤,2000)。
1.2.4 MDA和O2·含量测定MDA含量测定参照Heath等(1968)和Hodges等(1999)的方法,O2·含量测定参照王爱国等(1990)的方法。
1.2.5 叶绿素含量测定参照Arnon(1949)的方法。
1.2.6 总酚、类黄酮和谷胱甘肽含量测定总酚和类黄酮含量测定参照史国安等(2000)的方法,谷胱甘肽含量的测定参照中国科学院上海植物生理研究所等(1999)的方法。
2 结果 2.1 遮光对PAR及牡丹叶片Pn和Ci的影响由图 1可见,5月洛阳地区晴天中午自然光照强度最高可达2 310 μmol·m-2s-1。在遮光30%, 50%和80%处理条件下,一天之中最大光强分别为1 650,1 176和485 μmol·m-2s-1。

|
图 1 遮光对光合有效辐射日变化的影响 Figure 1 Effect of shading on PAR |
图 2显示,在不遮光处理条件下,牡丹叶片Pn的日变化有明显的“午休”现象,8:00—9:00 Pn处于最高水平(5.66 μmol·m-2s-1),随后开始下降,13:00左右降到最低(2.66 μmol·m-2s-1),以后又开始升高,15:00左右,出现了一个小峰值,之后又降低。几种遮光处理Pn都没有出现中午下降的现象,不遮光处理牡丹的Pn在中午最低时,遮光30%,50%和80%处理牡丹的Pn分别是不遮光的143%,115%和93%。在遮光30%处理条件下,Pn在一天中一直保持相对较高水平,在遮光50%次之。在遮光80%处理条件下,Pn在一天之中始终处于相对最低水平,16:00以后,Pn就出现了负值。

|
图 2 遮光对牡丹叶片Pn日变化的影响 Figure 2 Effect of shading on Pn |
在不遮光处理条件下,牡丹叶片Ci在8:00—14:30下降,在14:30—17:00迅速上升。在遮光30%处理条件下,Ci在8:00—13:00下降,13:00—17:00上升。在遮光50%处理条件下,Ci在8:00—11:00下降,11:00—17:00变化幅度很小。在遮光80%处理条件下,Ci的日变幅最大,从8:00—13:00快速下降,13:00—17:00又急剧升高(图 3)。

|
图 3 遮光对牡丹叶片Ci日变化的影响 Figure 3 Effect of shading on Ci |
图 4表明,在不遮光处理条件下,牡丹叶片Fv/Fm日变幅最大,从8:00—14:00,Fv/Fm下降很快,14:00时降至最低(0.805),14:00—18:00又迅速升高。在不同遮光处理条件下,中午Fv/Fm均呈下降趋势,但与不遮光处理相比,下降幅度较小,其中遮光80%处理的Fv/Fm变幅最小,其次为遮光50%,再次是遮光30%。

|
图 4 遮光对洛阳红Fv/Fm日变化的影响 Figure 4 Effect of shading on Fv/Fm |
遮光对牡丹叶片ФPSⅡ的影响与Fv/Fm相似(图 5)。不遮光处理,ФPSⅡ在12:00左右下降到最低。在不同遮光处理条件下,ΦPSⅡ的日变化趋势均是在7:00—12:00下降,12:00—18:00上升,3种遮光处理相比,12:00时,遮光80%处理的ΦPSⅡ下降最小,其次为遮光50%,再次是遮光30%。在遮光30%, 50%和80%处理条件下,ΦPSⅡ的最低值分别比不遮光处理的最低值提高69%, 106%和133%。

|
图 5 遮光对洛阳红ФPSⅡ日变化的影响 Figure 5 Effect of shading on ФPSⅡ |
由表 1可以看出,牡丹叶片的SOD活性以遮光30%处理最高(308.64 U·g-1 FW),分别比不遮光、遮光50%和遮光80%处理提高10.0%,11.1%和13.9%,差异达显著水平;牡丹叶片的POD活性以不遮光处理最高(56.04 U·min-1g-1 FW),分别比遮光30%、50%和80%处理提高25.9%,51.3%和181.2%。统计分析表明,除遮光30%和50%处理间牡丹叶片的POD活性差异不显著外,其他处理之间差异均达显著水平。
|
|

表 2表明,在遮光30%和50%处理条件下,叶片中MDA的含量显著低于不遮光处理,MDA含量分别比不遮光处理减少23.8%和19.3%,O2·的含量分别比不遮光处理减少15.2%和22.5%。在遮光80%处理条件下,MDA的含量比不遮光处理高11.4%, 而O2·的含量比不遮光处理降低24.1%。统计分析表明,MDA的含量遮光30%和50%与不遮光和遮光80%之间差异显著,O2·的含量不遮光与其他处理之间差异显著。
|
|
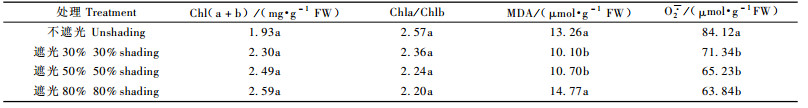
表 2显示,在不同遮光处理条件下,叶绿素总量及叶绿素b的相对含量都高于不遮光处理,并且随着遮光程度的增加,叶绿素总量及叶绿素b的相对含量增加,与不遮光处理相比,遮光30%,50%和80%处理的叶绿素总量分别增加19.2%,29.0%和34.2%,叶绿素b的相对含量分别增加29.3%,48.0%和57.3%。
2.7 遮光对牡丹叶片中总酚、类黄酮和谷胱甘肽含量的影响遮光显著影响牡丹叶片中总酚、类黄酮和谷胱甘肽的含量(表 3),遮光30%处理的总酚、类黄酮和谷胱甘肽含量均显著高于其他3个处理,总酚含量分别比不遮光、遮光50%和80%提高23.1%,17.6%和34.3%,类黄酮含量分别提高21.0%,35.9%和90.5%,谷胱甘肽含量分别提高75.0%,105.9%和250.0%。遮光80%处理的总酚、类黄酮和谷胱甘肽含量均显著低于其他3个处理。
|
|

研究结果显示,自然条件下的强光会影响牡丹的生长发育,过度遮光(遮光80%)同样对牡丹的生长有很大伤害,适度遮光则有利于牡丹生长发育,遮光30%与50%相比,在遮光30%处理条件下,牡丹的生长发育状况更好,主要体现在以下几个方面。
植物进行光合作用合成生命活动所需的各种物质,这是植物最根本的生命活动过程,Pn的大小直接关系到植物积累光合产物的快慢。本试验中,在不遮光处理条件下,在中午前后,牡丹叶片Pn日变化曲线上会出现一个谷值,而此时的光照强度达到一天之中的最高值,中午过后,随着光强的减弱,牡丹的Pn又有所升高,会出现一个小峰值,这与夏季很多植物的Pn在一天之中会出现“光合午休”的现象相似。在几种不同遮光处理条件下,Pn的变化都没有上述特点,它们的日变化曲线都是从8:00,9:00到13:00上升,13:00到17:00下降,按照Farquhar等(1982)的观点,光合速率的降低伴随着细胞间CO2浓度(Ci)的提高时,光合速率降低的主要原因一定是非气孔因素,是叶肉细胞光合活性的下降造成的。结合图 3可以看出Pn的降低与Ci的降低并不一致,Pn降低到最低时,Ci并没有降到最低,而Ci降到最低时,Pn却又有所升高。因此,可以推断,在不遮光处理条件下,Pn在中午降低可能是由于中午光照过强而非Ci的降低对牡丹产生了抑制作用。几种遮光处理相比,在遮光30%处理条件下,即使中午前后Ci有所降低,牡丹的Pn也始终保持在相对较高的水平,这有利于牡丹积累更多的光合产物,在遮光50%处理条件下,Pn介于遮光30%和遮光80%之间,在遮光80%处理条件下,牡丹的Pn一直很低,而且在16:00之后Pn就出现负值,这显然不利于牡丹积累光合产物。
Fv/Fm(PSⅡ最大光化学效率)反映ΦPSⅡ原初光能转化效率,非胁迫条件下该参数变化很小,不受物种和生长条件的影响,胁迫条件下该参数明显下降(寇伟锋等,2006),因此,它是反映植物光抑制程度的可靠指标,是光化学反应状况的重要参数。本研究表明,午间强光下,牡丹叶片Fv/Fm出现了不同程度的光抑制,但遮光处理的牡丹Fv/Fm的降低远没有不遮光处理显著,说明遮光处理减轻了牡丹叶片的光抑制程度。ΦPSⅡ表示用于电子传递的能量占PSⅡ所吸收光能的比例,ΦPSⅡ越高,就意味着PSⅡ光化学反应所利用的光能越多,同时表明过剩的光能越少,因此,光抑制程度越低;相反,则PSⅡ光化学反应所利用的光能越少,过剩的光能越多,光抑制程度越大(张守仁等,1999)。试验中,各遮光处理都提高了ΦPSⅡ,说明遮光处理使牡丹叶片对光能的利用率提高,光抑制程度降低。
SOD和POD在植物体内普遍存在,SOD是植物体内第一个清除活性氧的关键抗氧化酶, 它可以使Mehler反应中产生的O2·转化成H2O2, H2O2能进一步被CAT, POD所分解,从而使膜脂过氧化维持在一定水平。所以SOD和POD在自由基平衡中起重要作用,与植物的抗逆性和对环境的适应性密切相关(芦站根等,2003)。在不遮光处理条件下,虽然SOD和POD有相当高的活性,但同时MDA和O2·含量也高,证明SOD不足以清除所有的O2·,进而O2·对生物膜造成伤害产生MDA。在遮光30%处理条件下,SOD活性显著高于其他3个处理,虽然POD的活性不是最高,但结合表 2可以看出,此时,O2·含量低于不遮光处理,MDA含量在不同处理中最低,说明该处理牡丹受到较好的保护。在遮光80%处理条件下,虽然O2·含量较低,但SOD和POD活性最低,MDA含量最高,表明过度遮光对牡丹生长是不利的。
酚类物质、类黄酮和谷胱甘肽是植物体内的抗氧化剂,Maisuthisakul等(2007)认为植物体内的抗氧化活性主要是由酚类物质提供的,酚类物质与抗氧化活性直接相关,陈玉霞等(2007)的研究结果发现蔬菜的抗氧化活性与总多酚、总类黄酮含量间相关性较大。本研究发现,在遮光30%处理条件下总酚、类黄酮和谷胱甘肽含量在各个处理中最高,而且差异显著,因此,可以推断,遮光30%可能为牡丹创造了一个较适宜的生长环境,而遮光50%和80%则可能已经达到过度遮光的程度。
叶绿素是植物进行光合作用的物质基础,叶绿素包括叶绿素a和叶绿素b,前者主要利用长波光而后者主要吸收短波光进行光合作用。试验结果表明,遮光增加叶绿素总量和叶绿素b的相对含量,说明,遮光有利于牡丹利用更多的光能,也提高它对短波光的利用能力。
综上所述,洛阳地区的自然光照在中午前后对牡丹生长有一定的负效应,遮光能够改变牡丹的生长特性。几种不同程度遮光处理相比,在遮光30%处理条件下,牡丹的平均Pn在一天之中较高,Fv/Fm和ΦPSⅡ在中午左右下降较小,MDA和含量最低,总酚、类黄铜和谷胱甘肽含量最高,这些现象表明,遮光30%最有利于牡丹生长,本结果对进一步研究牡丹的光合生理以及对牡丹的栽培具有一定的参考价值。
陈玉霞, 郭长江, 杨继军, 等. 2007. 常见25种蔬菜抗氧化活性的比较研究[J]. 河南工业大学学报:自然科学版, 28(5): 37-41. |
高俊凤. 2000. 植物生理学实验技术[M]. 西安: 世界图书出版西安公司: 192-199.
|
寇伟锋, 刘兆普, 陈铭达, 等. 2006. 不同浓度海水对油葵幼苗光合作用和叶绿素荧光特性的影响[J]. 西北植物学报, 26(1): 73-77. |
刘卫琴, 汪良驹, 刘晖, 等. 2006. 遮荫对丰香草莓光合作用及叶绿素荧光特性的影响[J]. 果树学报, 23(2): 209-213. |
芦站根, 赵昌琼, 周文杰, 等. 2003. 光强对曼地亚红豆杉膜代谢及保护系统的影响[J]. 重庆大学学报, 26(8): 89-92. DOI:10.11835/j.issn.1000-582X.2003.08.024 |
史国安, 郭香凤. 2000. 槲叶加工过程中色素和酚类物质的变化[J]. 特产研究, (2): 14-16. |
王爱国, 罗广华. 1990. 植物的超氧物自由基与羟胺的定量关系[J]. 植物生理学通讯, 26(6): 55-57. |
王宪泽. 2002. 生物化学实验原理和方法[M]. 北京: 中国农业出版社: 186-190.
|
张国斌, 郁继华, 许耀照, 等. 2004. 低温弱光对辣椒幼苗叶绿素a荧光参数的影响[J]. 甘肃农业大学学报, 39(6): 614-619. |
张守仁. 1999. 叶绿素荧光动力学参数的意义及讨论[J]. 植物学通报, 16(4): 444-448. |
中国科学院上海植物生理研究所, 上海市植物生理学会. 1999. 现代植物生理学实验指南[M]. 北京: 科学出版社.
|
周治国, 孟亚利, 施培. 2001. 苗期遮荫对棉苗茎叶结构及功能叶光合性能的影响[J]. 中国农业科学, 34(5): 465-468. |
Arnon D J. 1949. Copper enzymes in isolated chloroplasts[J]. Plant Physiol, 24: 1-15. DOI:10.1104/pp.24.1.1 |
Demmig-Adams B, Adams W W Ⅲ. 1992. The xanthophyll cycle//Young A[J]. Carotenoids in photosynthesis. Berlin/Heidelberg: Springer: 405-432. |
Farquhar G D, Sharkey T D. 1982. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Physiology, 33: 317-345. DOI:10.1146/annurev.pp.33.060182.001533 |
Heath R L, Packer L. 1968. Photo-peroxidation in isolated chloroplasts: 1[J]. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophy, 125: 189-198. |
Hodges D M, Delong J M, Forney C F, et al. 1999. Improving the thiobarbituric acid -reactive -substances assay for estimating lipid peroxidation in plant tissues containing anthocynin and other interfering compounds[J]. Planta, 207: 604-611. DOI:10.1007/s004250050524 |
Maisuthisakul P, Pongsawatmanit R, Gordon M H. 2007. Characterization of the phytochemicals and antioxidant properties of extracts from team (Cratoxylum formosum Dyer)[J]. Food Chem, 100: 1620-1629. DOI:10.1016/j.foodchem.2005.12.044 |