 2010, Vol. 46
2010, Vol. 46文章信息
- 臧润国, 井学辉, 丁易, 成克武, 白志强, 张新平, 郭仲军, 张炜银
- Zang Runguo, Jing Xuehui, Ding Yi, Cheng Kewu, Bai Zhiqiang, Zhang Xinping, Guo Zhongjun, Zhang Weiyin
- 新疆阿尔泰山小东沟林区木本植物群落数量分类、排序及其环境解释
- Quantitative Classification, Ordination and Environmental Analysis of Woody Plant Communities in Xiaodonggou Forest Area of the Altai Mountain, Xinjiang
- 林业科学, 2010, 46(2): 24-31.
- Scientia Silvae Sinicae, 2010, 46(2): 24-31.
-
文章历史
- 收稿日期:2008-03-03
-
作者相关文章


2. 河北省承德市环境保护局 承德 067000;
3. 河北农业大学园林与旅游学院 保定 071000;
4. 新疆林业科学研究院森林生态研究所 乌鲁木齐 830000
2. Chengde Bureau of Environmental Protection Chengde 067000;
3. College of Landscape Architecture and Tourism, Agricultural University of Hebei Baoding 071000;
4. Institute of Forest Ecology, Xinjiang Academy of Forestry Science Urumuqi 830000
植物生态学的主要研究目标之一就是了解控制物种和群落分布的主要影响因子(Barton, 1993)。关于生物群落的异质性有2个经典学说,即环境控制论(Whittaker, 1956; Webb et al., 2000)和生物控制论(Schoener, 1983)。有研究表明,温度是控制海拔梯度上物种分布的主要限制因子(Daubenmire, 1943),也有人认为决定物种分布格局形成的因素取决于物种定居和灭亡的平衡(Münzbergová, 2004)。现代生态学家更加系统地分析环境因素与植被群落分布的关系,并认为干扰是导致物种镶嵌分布格局的重要因素(Wimberly et al., 2001)。
关于地形对植被格局的影响已开展大量的研究(Callaway et al., 1987; Thomas et al., 1993)。土壤因子对森林植被分布格局有不同尺度的影响(Johnston, 1992; Chen et al., 1997; Clark et al., 1999; Bigelow et al., 2002; John et al., 2007)。除以森林植被为研究对象外,也有学者对草地(Nelson et al., 1983)、荒漠植被(Xu et al., 2006)、河岸植被(Lenssen et al., 2005)的分布格局做了相关研究。
20世纪70年代, 随着计算机的诞生,多元统计方法如聚类分析和排序在生态学中得到广泛的应用。20世纪80年代作为间接梯度分析的DCA排序方法被广泛应用于生态学研究(HillandGauch, 1980),但该方法没有涉及环境数据,而是间接从物种组成数据来推算环境因子对物种分布格局的影响。近年来,作为直接梯度分析的CCA和DCCA因其能同时结合多个环境因子、包含大量的信息、结果准确等优点得到广泛应用(AllenandPeet, 1990; Odland et al., 1990)。目前,应用于物种-环境关系研究最流行方法之一是去趋势典范对应分析(Detrended canonical correspondence analysis, DCCA),它是一种限定性排序方法,即在每一轮样方值-物种值的加权平均叠代运算后,用环境因子和样方排序值做一次多元线性回归,用回归系数与环境因子原始值计算出样方分值,再进行新一轮叠代计算,由此得出的排序轴代表环境因子的一种线性组合,消除了“弓形效应”,因此成为现代植被梯度分析与环境解释的流行方法(Zhang et al., 1994; 邱扬等, 2000; 沈泽昊, 2002; Xu et al., 2006)。
阿尔泰山是亚洲的宏大山系之一,东起蒙古巴彦温都尔,经新疆北部向西延伸至俄罗斯鲁布佐夫斯克,横亘蒙古、中国、哈萨克斯坦、俄罗斯四国,长约1 650 km,宽约300 km,呈西北—东南走向。中国境内的阿尔泰山属于其中段的西南坡,东西长约450 km,南北宽约80~150 km,呈西北高而宽,东南低而窄的地形特征。阿尔泰山由一系列南西向北东逐渐升高的阶梯状山地组成,切割强烈。一般海拔在1 000~3 500 m,其中北部最高的友谊峰海拔4 374 m。本文所指的阿尔泰山山地植被指分布于我国境内阿尔泰山南坡中段的小东沟林区辖域范围内的植被。小东沟林区山地植被随地貌和海拔变化,呈现明显的带状更替现象。山地森林主要分布在海拔1 300~2 600 m,阴坡林分郁闭度明显高于阳坡。不同森林群落类型分布具有明显的界线。
目前国内已经对阿尔泰山林区的植被开展了一些研究,主要集中在森林资源动态变化(时旭辉等, 1995)、种子植物区系(陈文俐等, 2000)、树种天然更新特点及规律(曾东等, 2000)及森林类型划分(臧润国等, 2005)。这些研究多数是定性描述的,关于该区物种梯度分布与环境解释的定量研究还未见报道。
本文利用野外调查的群落样地资料,对小东沟林区植物群落类型进行了划分,并利用去趋势典范对应分析(DCCA)方法,探讨了小东沟林区物种分布与环境因子的关系,旨在揭示小东沟林区主要植物群落分布格局及影响因子,以期为今后在阿尔泰林区开展森林生态系统功能、生物多样性保育和林区资源可持续发展等方面的研究和相应的经营管理提供科学依据。
1 研究区概况阿勒泰地区的地理位置为45°59′—49°10′ N,85°31′—91°01′ E。属于大陆性寒温带气候。气候多严寒,冷暖悬殊,年较差大,日较差也显著。年均温4.5 ℃,1月平均气温-16.7 ℃,7月平均气温22.1 ℃,极端最低气温-43.5 ℃,极端最高温度37.6 ℃。冬季寒冷期>110天,严寒期(≤-20 ℃)约58~63天。年均降水量183 mm,主要集中在6—12月,占年降水量的70%~80%。5—6月山地融雪水与降雨叠加容易形成洪水。年均日照时数3 010.8 h。年均风速1.0~5.0 m·s-1,多8级以上大风,春季最多,夏季次之,秋冬天气比较稳定。年降雪140~150天,积雪深度50~60 cm,最深可达89 cm。
研究区位于小东沟森林公园内的小东沟山系,距离阿勒泰市区北约10 km处。年均温度-4~-2 ℃。年降水较丰富,且降水量随海拔的升高而递增,同时也从北向南、从西向东逐渐减少。年降水量中山带300~500 mm,高山带600~800 mm。夏季多雨,冬季积雪,气候、地貌和植被垂直带分布明显。林区主要土壤有山地栗钙土和山地灰色森林土。该林区乔木树种比较单纯,主要有新疆落叶松(Larix sibirica),还有较少的新疆云杉(Picea obovata)、新疆冷杉(Abies sibirica),落叶阔叶树种主要有欧洲山杨(Populus tremula)、疣枝桦(Betula pendula)和苦杨(Populus laurifolia)等。灌丛种类丰富,有多种刺柏属(Juniperus)、圆柏属(Sabina)、蔷薇属(Rosa)、栒子属(Cotoneaster)、忍冬属(Lonicera)、小檗属(Berberis)、绣线菊属(Spiraea)等种类。
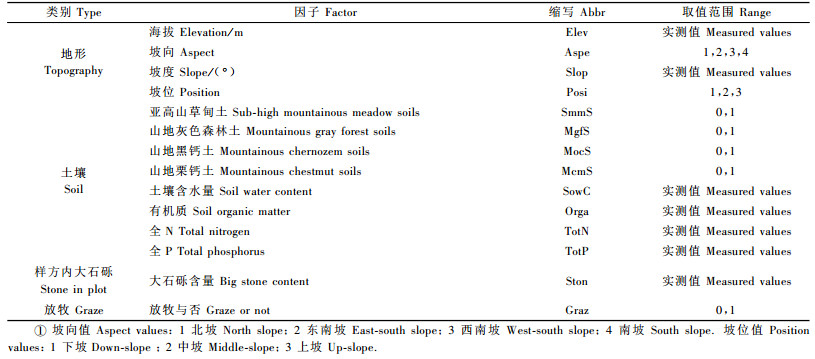
2 研究方法 2.1 调查方法为了反映整个垂直梯度带上的物种分布格局,分别在小东沟南、北两坡向沿海拔1 100~1 800 m和1 200~2 200 m设置调查样带。以每50 m的海拔为间隔设置一个20 m×20 m的样地,并将其分成16个5 m×5 m的小样方调查其乔木和灌木。共调查样地43个,其中南、北坡分别为18个和25个。调查内容包括: 1)群落特征:在每个样地内用样绳分成5 m×5 m的小样方,在每个小样方内记录DBH≥1 cm的乔木种名、胸径和树高。对DBH < 1 cm的乔木幼树和幼苗按高度≤0.5 m和>0.5 m 2类统计各种的数量。小样方内的灌木记录种名、高度和盖度。2)环境因子:利用GPS在样地中心位置记录样地的地理坐标和海拔,用罗盘实测样地坡度,同时记录样地所在坡向、坡位、样方内大石砾含量及受放牧干扰与否。3)土壤:群落调查结束后,在每一样地内选择代表平均状况的地点挖掘一土壤剖面,用铝盒取0~20 cm土层中的土样,秤土样质量,另取适量土放入土袋,带回实验室进行分析。土样分析指标包括有机质、全N和全P含量。每样地秤过质量的铝盒样品带回室内再经过48 h、105 ℃烘干,待铝盒冷却秤量土样干质量,计算样地土壤含水量。
2.2 数据预处理及其变量确定根据各样地内物种的多度、频度等,分别计算乔木、灌木层的重要值,其中样地内的幼苗和幼树都归为灌木层统计。计算公式如下:

|
为了满足DCCA数据分析的需要,对于定量变量,如海拔、坡度等数据,采用实测值表示。对于虚变量采用0,1赋值的形式,如以样地是否受放牧干扰为例,样地受放牧影响赋值为1,不受放牧影响赋值为0。对于定性的有序多变量数据,采用分级方法赋值,如坡向(ASPE)从阴坡到阳坡共划分4级,分别赋值1~4,数值增加表明样地光照强度增加,越向阳,越干热。坡位(POSI)划分下坡、中坡和上坡,分别赋值1~3。
在进行DCCA分析之前,为了消除各因子取值范围与量纲不同的影响,并满足变量数据的正态要求,对坡度、海拔、大石砾含量、土壤含水量数据进行标准化转换X′i=(Xi-Xmin)/(Xmax-Xmin)。对土壤全N、全P和有机质数据做了对数转换,以更好地区分微小数值变化对植被分布格局的影响。
2.3 群落类型划分和排序应用SPSS 13.0软件的聚类分析方法对调查的43个样地进行类型划分。首先确定聚类分析所用的数据集,即统计各样方的乔木层和灌木层物种的重要值。其次,确定最多划分水平数量,其余按照系统默认程序进行。本研究的物种-环境因子排序采用DCCA方法,它是结合趋势对应分析(DCA)与典范对应分析(CCA)发展起来的,因其排序轴既包含物种信息,又包含环境信息,又克服弓形效应,是目前国际上最新的排序方法(邱扬等, 2000; Xu et al., 2006)。它可以直观地把环境因子、物种、样方同时表达在排序轴的坐标平面上。分析采用CANOCO for windows, Version 4.5软件,所应用的环境因子为14个因子(表 1),物种数据采用各样地内物种的重要值。对物种和环境数据做DCCA分析时,为了满足软件的需要,对物种重要值数据做了对数转换,其他均按照系统默认选项进行。
|
|
在调查的43个样地内,共记录23个乔灌物种,分属于16属和14科。通过聚类对小东沟林区43个样地进行6个群落类型的划分(表 2)。
|
|
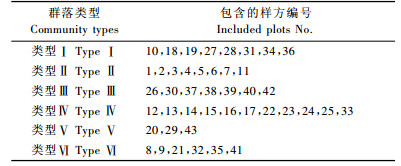
群落类型Ⅰ是以欧洲山杨为主的混交阔叶林,多与疣枝桦混生分布,也有少量的新疆云杉出现,林下灌木层主要有绣线菊、腺齿蔷薇(Rosa albertii)、臭茶藨(Ribes graveolens)等。群落类型Ⅱ主要是以新疆落叶松为优势种的针叶林,混有少量的新疆云杉和新疆冷杉,其林下灌木层优势种为新疆圆柏(Sabina vulgalis),腺齿蔷薇少有分布。群落类型Ⅲ主要是灌木群落类型,主要分布在阳坡,优势种是各种绣线菊和忍冬,亦有新疆圆柏、小檗、接骨木(Sambucus williamsii)等灌木伴生出现。群落类型Ⅳ主要是针叶混交林,优势种有新疆冷杉、新疆落叶松和新疆云杉,有时伴有阔叶树种疣枝桦出现。林下灌木层的优势种主要是乔木幼苗,包括新疆冷杉、新疆云杉幼苗,灌木树种忍冬、腺齿蔷薇等少有分布。群落类型Ⅴ主要是以苦杨为优势种的阔叶林,常与疣枝桦混生分布。林下灌木层优势种主要是苦杨幼苗,臭茶藨灌木少有分布。群落类型Ⅵ是以疣枝桦为优势种的混交群落类型,主要与黄花柳(Salix caprea)混交,有时混有少量的新疆落叶松、新疆云杉等针叶树种。林下灌木层以乔木幼苗或腺齿蔷薇占优势,伴有臭茶藨、接骨木等混生分布。
3.2 环境因子之间的相关性分析不同环境因子之间存在显著的相关性(表 3),海拔和山地灰色森林土土壤类型与坡位均呈极显著的正相关,相关系数分别为0.933和0.658。山地栗钙土土壤类型、土壤全N和土壤全P与坡位呈极显著负相关,相关系数分别为-0.923, -0.477和-0.602。亚高山草甸土土壤类型和山地灰色森林土土壤类型与海拔均呈极显著的正相关,相关系数分别为0.441和0.602,山地栗钙土土壤类型、土壤全N和土壤全P与海拔均呈极显著的负相关,相关系数分别为-0.860, -0.456和-0.603。放牧与坡度呈显著的负相关,相关系数为-0.327。
|
|
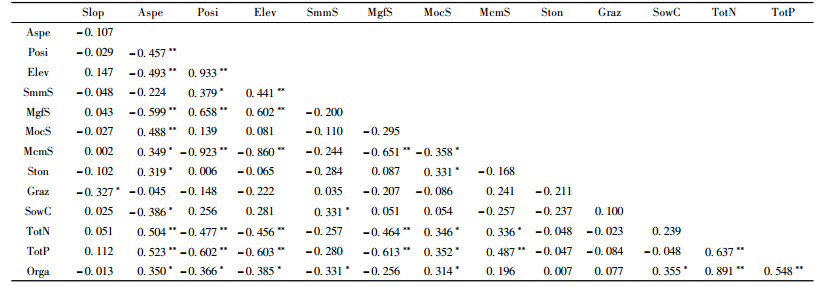
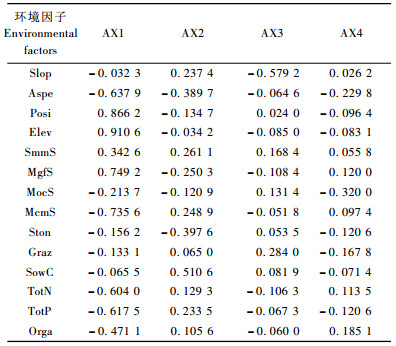
通过DCCA排序,14个环境因子中与轴1相关性最高的环境因子是海拔,相关系数为0.910 6;其次是坡位,相关系数为0.866 2;山地灰色森林土土壤类型、山地栗钙土土壤类型、土壤全P、土壤全N、坡向均与DCCA的第1轴具有明显的相关性。其中,山地栗钙土土壤类型、土壤全P、全N、坡向这4个环境因子与DCCA第1轴呈负相关(相关系数分别为-0.735 6, -0.617 5, -0.604 0, -0.637 9)。与第2轴相关性最高的环境因子是土壤含水量,相关系数为0.510 6。坡向和样地内的大石砾含量与DCCA第2轴呈负相关,相关系数分别为-0.389 7和-0.397 6。14个环境因子中,与DCCA第3轴相关性最高的环境因子是坡度因子,相关系数为-0.579 2。与第4轴相关性较高的环境因子是山地黑钙土土壤类型,呈负相关性。
|
|
不同的群落类型沿着相应环境梯度具有不同的分布范围(图 1)。群落类型Ⅰ主要分布在DCCA排序图的左边,一般海拔介于1 200~1 700 m,坡度在20~30°。阴坡和阳坡均有分布,从阴坡到阳坡,样地内大石砾含量显著增加,少数样地内大石砾含量可达到90%。沿海拔梯度,土壤类型由山地栗钙土向山地黑钙土转化。坡度陡峭的地方,不受放牧的干扰。群落土壤养分条件相对较好。

|
图 1 样方和环境因子DCCA排序 Figure 1 DCCA result for ordination of plots and environmental factors |
群落类型Ⅱ主要分布于DCCA图的第2象限,一般海拔分布范围2 000~2 200 m,属于高海拔地段,个别分布在海拔1 500 m左右。该类型群落主要分布在阴坡,坡位较高处,坡度较缓,少数地势陡峭。土壤类型海拔2 100 m以下以山地灰色森林土为主,海拔2 100 m以上亚高山草甸土广泛分布。样地内的大石砾含量明显减少。土壤水分条件好,土壤全N、全P、有机质含量明显小于群落类型Ⅰ。坡度小的地方仍受放牧影响。
群落类型Ⅲ主要分布在阳坡、坡位较低的地带。物种主要以灌木为主。一般分布在海拔1 200~1 400 m,个别出现在海拔1 800 m左右,由于海拔分布低,牲畜容易接近,受放牧影响严重。样地内土壤相对较干,土壤类型趋向于山地栗钙土。
群落类型Ⅳ主要分布在DCCA图的第4象限,该群落类型主要分布在阴坡、坡度平缓处,坡度在13~25°,海拔1 600~2 000 m,群落多分布在中坡、坡顶位置。群落内的大石砾含量介于20%~40%之间。受放牧影响。土壤类型主要为山地灰色森林土。样地内的土壤养分相对较肥沃, 土壤湿润。
群落类型Ⅴ主要分布在DCCA排序图的左侧,处于坡度平缓的河谷地带,海拔1 100~1 300 m,牲畜容易接近,严重受放牧干扰。主要生长以苦杨为优势种的阔叶混交林群落类型,土壤类型以山地栗钙土为主。土壤水分条件好,土壤养分条件也优于群落类型Ⅱ和Ⅳ。
群落类型Ⅵ主要是以疣枝桦为优势种的阔叶混交群落类型,该群落类型在DCCA排序图上位于左边,多分布在下坡位,坡度平缓,个别地段较陡。海拔范围1 100~1 500 m。土壤类型以山地栗钙土为主,山地黑钙土也有少量分布。样地土壤养分中以全P含量较高,土壤全N、有机质含量相对较低。
不同植物种类对环境条件的需求存在差异,因而具有不同的分布范围(图 2,由于乔木层的幼苗放到灌木层统计,所以图中将每种乔木幼苗按照一个物种参与排序,其中含7种乔木的幼苗)。DCCA排序图第2象限内的物种主要是新疆落叶松,林下灌木为新疆圆柏,其分布海拔相对较高。土壤类型为山地灰色森林土。第4象限内的物种主要沿着海拔、坡位和山地灰色森林土土壤类型3个环境因子分布,主要乔木种类为新疆冷杉和新疆云杉。象限1内主要分布的物种为阔叶树种,包括乔木树种欧洲山杨、疣枝桦、苦杨、黄花柳,林下灌木层除了乔木幼苗,还有高山绣线菊。象限3内主要分布的物种为灌木种类,主要包括腺齿蔷薇、金丝桃叶绣线菊(Spiraea hypericifolia)、欧亚绣线菊(Spiraea media)、臭茶藨、小叶忍冬(Lonicera nigra)、黑果忍冬(Lonicera microphylla)等。这类灌木群落多分布于中低海拔、光照充足、土壤干燥的环境中,但受放牧影响很大。象限2和象限4内的物种分布海拔范围比象限1和象限3高,分布所处的坡位值也大,且一般分布针叶林,多分布在阴坡。象限1和象限3内多分布阔叶乔木树种和灌丛,多分布在中低海拔、光照强、土壤养分含量相对较高的环境。

|
图 2 物种和环境因子DCCA排序 Figure 2 DCCA result for ordination of species and environmental factors 物种名 Species name : 1 新疆落叶松 Larix sibirica; 2 新疆冷杉 Abies sibirica; 3 新疆云杉 Picea obovata; 4 黄花柳 Salix caprea; 5 疣枝桦 Betula pendula; 6 欧洲山杨 Populus tremula; 7 苦杨 Populus laurifolia; 8 新疆落叶松幼苗 Larix sibirica seedlings; 9 新疆圆柏 Sabina vulgalis; 10 新疆冷杉幼苗 Abies sibirica seedlings; 11 新疆云杉幼苗 Picea obovata seedlings; 12 黑果栒子 Cotoneaster melanocarpus; 13 疣枝桦幼苗 Betula pendula seedlings; 14 腺齿蔷薇 Rosa albertii; 15 高山绣线菊 Spiraea alpina; 16 新疆忍冬 Lonicera tatarica; 17 欧洲山杨幼苗 Populus tremula seedlings; 18 欧洲刺柏 Juniperus formosana; 19 臭茶藨 Ribes graveolens; 20 苦杨幼苗 Populus laurifolia seedlings; 21 黑果小檗 Berberis atrocarpa; 22 石生悬钩子 Rubus saxatilis; 23 大叶绣线菊 Spiraea chamaedryfolia; 24 接骨木 Sambucus williamsii; 25 树莓 Rubus swinhoei; 26 金丝桃叶绣线菊 Spiraea hypericifolia; 27 欧亚绣线菊 Spiraea media; 28 小叶忍冬 Lonicera nigra; 29 黑果忍冬 Lonicera microphylla; 30 黄花柳幼苗 Salix caprea seedlings; 31 小叶茶藨 Ribes heterotrichum. |
数量分类和排序是研究植被与环境关系的基本方法(Mucina, 1997)。目前应用于植被分类的方法有二元指示种分类法(TWINSPAN)(Ase et al., 1993; Fensham, 1995; Xu et al., 2006)、聚类法(cluster analysis)(刘世梁等, 2003)和依据群落外貌与种类组成相结合的划分方法(宋创业等, 2007)等。本研究采用聚类分析将43个样地分成6个群落类型,其中包括5个森林类型,1个灌丛类型,分别为欧洲山杨林、新疆落叶松林、灌丛群落、新疆冷杉+新疆落叶松+新疆云杉林、苦杨林和疣枝桦林,数量分类的群落类型与野外调查观察的类型非常一致。
DCCA因其具有许多优点,是目前较新的排序方法,被广泛运用于多种植物群落研究(邱扬等, 2000; 张峰等, 2003; Zhao et al., 2005; Xu et al., 2006)。DCCA排序同时结合物种和环境因子信息,因此能较好表达群落的环境梯度,反映环境的空间变异特点。第1排序轴主要表达寒冷、土壤贫瘠方向的梯度,主要反映海拔、坡位等方面的变异。第2轴主要表达群落沿土壤水分梯度的变化。6类群落类型分布的生境各有特点。第Ⅰ群落类型在阴坡主要分布在低海拔,在阳坡主要分布在中高海拔,且样地内石砾含量较高。第Ⅱ群落类型主要分布在阴坡高海拔,生境具有寒冷、干旱、土壤贫瘠的特点。第Ⅲ群落为灌丛类型,主要分布在阳坡低海拔,地势平坦,生境暖干。第Ⅳ群落类型为针叶混交林,主要分布在阴坡中高海拔,海拔分布仅低于群落类型Ⅱ,生境特点冷湿。第Ⅴ群落类型主要分布在低海拔、地势平坦的河谷地段,土壤水分条件好。第Ⅵ群落类型主要在阴坡、半阳坡的中低海拔,地势相对平坦处。
大量研究表明,在区域至全球尺度上,气候条件是决定植物类型或生活型分布的主要因素(Woodward et al., 1991)。在景观、群落或更小尺度上,地形和土壤这种非地带性的环境因子是影响物种分布格局的主要因素(Henriques et al., 1998; Clack et al., 1999)。山区森林植被格局通常认为海拔变化是控制物种群落组成最主要的因素,海拔升高,导致温度、降水、地形有所不同,从而影响土壤湿度和光照条件,进而影响物种分布和个体生长(Johnson et al., 1968; del Moral et al., 1978; Wimberly et al., 2001; Zhao et al., 2005)。研究证实,温度和降水以及所有能改变温度和降水的因素对自然植被分布均有影响(Zhao et al., 2005)。本研究结果表明,与DCCA排序第1轴相关性最高的是海拔,其次是坡位、土壤类型、土壤养分等因子; 与第2轴相关性最高的环境因子是土壤水分。总的来看,地形因子是影响小东沟山区植被分布的最主要环境因子,其次是土壤因子,即地形和土壤因子综合作用影响阿尔泰山小东沟山区植被分布。这和多数山区森林植被-环境关系研究得出的结论相一致(Allen et al., 1991; Oliveira-Filho et al., 1998; 张金屯, 2005; 刘秋锋等, 2006)。
然而,也有研究认为自然和人为干扰是形成目前物种分布格局的最主要影响因子(Hadley, 1994; Thompson et al., 2002),而且能够显著影响垂直梯度上的物种变化格局(Nogués-Bravo et al., 2008)。虽然与长期的气候变化和地质运动对植被影响相比,人类干扰历史相对较短,但是对自然生态系统造成严重影响(Zhao et al., 2005)。阿勒泰地区是新疆乃至全国的重点牧区之一,每年5—6月山地融雪水与降雨叠加形成洪水时,额尔齐斯河两岸大面积被水淹没,牧民将放牧场所转移到山区,从而不断造成林牧矛盾。为了促进畜牧业的发展,当地牧业管理部门允许牧民放牧,并颁发了长期使用的草场证。牲畜不仅在山区草地啃食,同时也常常进入林地内啃食。牲畜的啃食直接破坏幼苗幼树,导致森林树木无法正常更新。过大的牲畜密度也对草原生态系统产生严重危害,草地生态系统发生不同程度的退化。有些物种由于受牲畜过度啃食和践踏,更新困难,从而改变了山区物种的分布格局。在阿尔泰林区,长期放牧已经导致各种森林群落类型的组成和结构的重大变化,树木难以更新,生态系统的多种服务功能难以正常发挥,但放牧干扰目前还没有对不同群落类型的空间分布格局造成显著影响,各主要群落类型仍然按照其在不同的海拔、坡向或土壤梯度占据着空间上相应的生态位。放牧在一定程度上影响了不同群落类型分布的边界,但从野外调查来看,各类型的空间面积没有显著变化。在分析阿尔泰林区的森林群落类型分布时,还应该考虑采伐干扰,因为采伐也可能明显改变不同类型群落的结构,甚至在大强度采伐后群落类型可能会发生改变,如云杉或落叶松林就很有可能被欧洲山杨林所取代,有关采伐对阿尔泰林区群落类型及其分布格局的影响还需要从景观动态的角度作进一步的细致研究。所调查研究的小东沟林区所经历的采伐并不多,因此其群落类型及其分布格局基本上代表了这一区域森林植被类型的自然格局,地形和土壤是控制植被格局的关键因素。
陈文俐, 杨昌友. 2000. 中国阿尔泰山种子植物区系研究[J]. 云南植物研究, 22(4): 371-378. |
刘秋锋, 康慕谊, 刘全儒. 2006. 中条山混沟地区森林乔木种的数量分类与环境解释[J]. 植物生态学报, 30(3): 383-391. DOI:10.17521/cjpe.2006.0052 |
刘世梁, 马克明, 傅伯杰, 等. 2003. 北京东灵山地区地形土壤因子与植物群落关系研究[J]. 植物生态学报, 27(4): 496-502. DOI:10.17521/cjpe.2003.0072 |
邱扬, 张金屯. 2000. DCCA排序轴分类及其在关帝山八水沟植物群落生态梯度分析中的应用[J]. 生态学报, 20(2): 199-206. |
沈泽昊. 2002. 山地森林样带植被-环境关系的多尺度研究[J]. 生态学报, 22(4): 461-470. |
时旭辉, 周林生. 1995. 新疆天山北坡及阿尔泰山两大林区森林资源动态的研究[J]. 八一农学院学报, 18(2): 9-15. |
宋创业, 郭柯. 2007. 浑善达克沙地中部丘间低地植物群落分布与土壤环境关系[J]. 植物生态学报, 31(1): 40-49. |
臧润国, 成克武, 李俊清, 等. 2005. 天然林生物多样性保育与恢复[M]. 北京: 中国科学技术出版社.
|
曾东, 李行斌, 于恒. 2000. 新疆落叶松、新疆云杉迹地天然更新特点与规律的辨析[J]. 干旱区研究, 17(3): 46-52. |
张峰, 张金屯. 2003. 历山自然保护区猪尾沟森林群落植被格局及环境解释[J]. 生态学报, 23(3): 421-427. |
张金屯. 2005. 历山自然保护区森林群落的典范主分量分析[J]. 生物数学学报, 20(2): 213-218. |
Allen R B, Peet R K. 1990. Gradient analysis of forests of the Sangre de Cristo Range, Colorado[J]. Canadian Journal of Botany, 68: 193-201. DOI:10.1139/b90-026 |
Allen R B, Peet R K, Baker W L. 1991. Gradient analysis of latitudinal variation in southern Rocky mountain forests[J]. Journal of Biogeography, 18: 123-139. DOI:10.2307/2845287 |
Ase M, Birks H J B. 1993. A numerical analysis of the distribution patterns of Salix species in Europe[J]. Journal of Biogeography, 20: 1-32. DOI:10.2307/2845736 |
Barton A M. 1993. Factors controlling plant distributions: drought, competition, and fire in montane pines in Arizona[J]. Ecological Monographs, 63: 367-397. DOI:10.2307/2937151 |
Bigelow S W, Canham C D. 2002. Community organization of tree species along soil gradients in a north-eastern USA forest[J]. Journal of Ecology, 90: 188-200. DOI:10.1046/j.0022-0477.2001.00655.x |
Callaway R M, Clebsch E E C, White P S. 1987. A multivariate analysis of forest communities in the western Great Smoky Mountains National Park[J]. American Midland Naturalist, 118: 107-120. DOI:10.2307/2425633 |
Chen Z S, Hsieh C F, Jiang F Y, et al. 1997. Relation of soil properties to topography and vegetation in a subtropical rain forest in southern Taiwan[J]. Plant Ecology, 132: 229-241. DOI:10.1023/A:1009762704553 |
Clark D B, Palmer M W, Clark D A. 1999. Edaphic factors and the landscape-scale distributions of tropical rain forest trees[J]. Ecology, 80: 2662-2675. DOI:10.1890/0012-9658(1999)080[2662:EFATLS]2.0.CO;2 |
Daubenmire R F. 1943. Vegetation zonation in the Rocky Mountains[J]. Botanical Review, 9: 325-393. DOI:10.1007/BF02872481 |
Del Moral R, Watson A E. 1978. Gradient structure of forest vegetation in the Central Washington Cascades[J]. Vegetatio, 38: 29-48. DOI:10.1007/BF00141297 |
Fensham R J. 1995. Floristics and environmental relations of inland dry rainforest in North Queensland, Australia[J]. Journal of Biogeography, 22: 1047-1063. DOI:10.2307/2845834 |
Hadley K S. 1994. The role of disturbance, topography, and forest structure in the development of a montane forest landscape[J]. Bulletin of the Torrey Botanical Club, 121: 47-61. DOI:10.2307/2996883 |
Henriques R, Hay J. 1998. The plant communities of a foredune in southeastern Brazil[J]. Canadian Journal of Botany, 76: 1323-1330. |
Hill M O, Gauch H G. 1980. Detrended correspondence analysis: an improved ordination technique[J]. Vegetatio, 42: 47-58. DOI:10.1007/BF00048870 |
John R, Dalling J W, Harms K E, et al. 2007. Soil nutrients influence spatial distributions of tropical tree species[J]. Proceedings of the National Academy of Sciences, 104: 864-869. DOI:10.1073/pnas.0604666104 |
Johnson M, Mason L, Raven P. 1968. Ecological parameters and plant species diversity[J]. American Naturalist, 102: 297-306. DOI:10.1086/282544 |
Johnston M H. 1992. Soil-vegetation relationships in a Tabonuco forest community in the Luquillo mountains of Puerto Rico[J]. Journal of Tropical Ecology, 8: 253-263. DOI:10.1017/S0266467400006477 |
Lenssen J P M, De Kroon H. 2005. Abiotic constraints at the upper boundaries of two Rumex species on a freshwater flooding gradient[J]. Journal of Ecology, 93: 138-147. DOI:10.1111/jec.2005.93.issue-1 |
Münzbergová Z. 2004. Effect of spatial scale on factors limiting species distributions in dry grassland fragments[J]. Journal of Ecology, 92: 854-867. DOI:10.1111/jec.2004.92.issue-5 |
Mucina L. 1997. Classification of vegetation: past, present and future[J]. Journal of Vegetation Science, 8: 751-760. DOI:10.2307/3237019 |
Nelson D C, Anderson R C. 1983. Factors related to the distribution of prairie plants along a moisture gradient[J]. American Midland Naturalist, 109: 367-375. DOI:10.2307/2425417 |
Nogués-Bravo D, Araújo M B, Romdal T, et al. 2008. Scale effects and human impact on the elevational species richness gradients[J]. Nature, 453: 216-219. DOI:10.1038/nature06812 |
Odland A, Birks H J B, Line J M. 1990. Quantitative vegetation-environment relationships in west Norwegian tall-fern vegetation[J]. Nordic Journal of Botany, 10: 511-533. DOI:10.1111/j.1756-1051.1990.tb02095.x |
Oliveira-Filho A T, Curi N, Vilela E A, et al. 1998. Effects of canopy gaps, topography, and soils on the distribution of woody species in a central Brazilian deciduous dry forest[J]. Biotropica, 30: 362-375. DOI:10.1111/btp.1998.30.issue-3 |
Schoener T W. 1983. Field experiments on interspecific competition[J]. American Naturalist, 122: 240-285. DOI:10.1086/284133 |
Thomas R L, Anderson R C. 1993. Influence of topography on stand composition in a midwestern ravine forest[J]. American Midland Naturalist, 130: 1-12. DOI:10.2307/2426270 |
Thompson J, Brokaw N, Zimmerman J K, et al. 2002. Land use history, environment, and tree composition in a tropical forest[J]. Ecological Applications, 12: 1344-1363. DOI:10.1890/1051-0761(2002)012[1344:LUHEAT]2.0.CO;2 |
Webb C, Peart D. 2000. Habitat associations of trees and seedlings in a Bornean rain forests[J]. Journal of Ecology, 88: 464-478. DOI:10.1046/j.1365-2745.2000.00462.x |
Whittaker R H. 1956. Vegetation of the Great Smoky Mountains[J]. Ecological Monographs, 26: 1-80. DOI:10.2307/1943577 |
Wimberly M C, Spies T A. 2001. Influences of environment and disturbance on forest patterns in coastal Oregon watersheds[J]. Ecology, 82: 1443-1459. DOI:10.1890/0012-9658(2001)082[1443:IOEADO]2.0.CO;2 |
Woodward F I, Mckoo I F. 1991. Vegetation and climate[J]. Environment International, 17: 535-546. DOI:10.1016/0160-4120(91)90166-N |
Xu L, Liu H, Chu X, et al. 2006. Desert vegetation patterns at the northern foot of Tianshan Mountains: the role of soil conditions[J]. Flora, 201: 44-50. DOI:10.1016/j.flora.2005.04.003 |
Zhang J T, Oxley E R B. 1994. A comparison of three methods of multivariate analysis of upland grasslands in north Wales[J]. Journal of Vegetation Science, 5: 71-76. DOI:10.2307/3235640 |
Zhao C M, Chen W L, Tian Z Q, et al. 2005. Altitudinal pattern of plants species diversity in Shennongjia Mountain, central China[J]. Journal of Integrative Plant Biology, 47: 1431-1449. DOI:10.1111/jipb.2005.47.issue-12 |