 2009, Vol. 45
2009, Vol. 45文章信息
- 周光益, 田大伦, 杨乐苏, 王旭, 邱治军, 刘敏.
- Zhou Guangyi, Tian Dalun, Yang Lesu, Wang Xu, Qiu Zhijun, Liu Min
- 酸雨胁迫下乡土植物根际土壤的化学行为
- Chemical Behavior of Rhizosphere Soil of Native Plants to Different Acid Rain Stress
- 林业科学, 2009, 45(11): 161-166.
- Scientia Silvae Sinicae, 2009, 45(11): 161-166.
-
文章历史
- 收稿日期:2009-03-06
-
作者相关文章


2. 中国林业科学研究院热带林业研究所 广州 510520
2. Research Institute of Tropical Forestry, CAF Guangzhou 510520
根际是指围绕植物活根周围的土壤微域,由于受根系生物活动的影响,在物理、化学和生物特性上不同于原土体。本文采用Riley等(1971)的方法来区分根际土与非根际土。酸雨作为外来酸进入森林土壤后,必然对土壤的物理化学、生物化学等性质产生影响。许多研究者从不同角度分析了酸雨对土壤的影响(Hagedorn et al., 2001; 岑慧贤等, 2001; 樊后保等, 2002; Delfosse et al., 2004; 高太忠等, 2004);但是从酸雨胁迫下植物根际土壤化学响应方面的研究较少(Samuels et al., 1997),目前普遍认为酸雨对植物的伤害一方面酸雨淋溶对植物地上部分的直接伤害,另一重要方面酸雨在导致土壤酸化过程中对植物的间接影响,而植物与土壤的作用中,根际土起关键作用,决定着植物实际吸收的养分量,与生长关系密切(罗微等,2004),且根际土与非根际土的化学行为是有区别的。为此,本研究从利用短期暴露在不同酸雨污染环境下的植物、长期在不同酸沉降胁迫环境的植物,分析植物根际和非根际土壤酸雨胁迫下土壤pH和有效养分的变化规律以及有害物质(酸物质及Al3+)的富集情况,阐述不同酸胁迫下森林土壤的化学行为和响应,为分析酸雨对土壤及植物影响提供参考。
1 材料与方法 1.1 植物短期暴露试验及相关样品的采集1) 植物材料:在代表污染区的广州市黄埔区广州石化厂后山(酸雨频率达95%以上)和代表相对清洁区的从化流溪河国家森林公园(酸雨频率达69.5%)开展植物短期暴露的盆栽试验,选取热带和南亚热带森林中常见的乡土植物种19种,包括山杜英(Elaeocarpus sylvestris)、土沉香(Artocarpus hypargyreus)、大头茶(Gordonia axillaris)、乌墨(Syzygium cumini)、荷木(Schima superba)、小叶榄仁(Terminalia mantaly)、幌伞枫(Heteropanax fragrans)、雄黄豆(Cassia javanica)、灰木莲(Manglietia glauca)、阴香(Cinnamomum burmanni)、火力楠(Michelia macclurei)、樟树(Cinnamum camphora)、孔雀豆(Adenanthera paeonina)、秋枫(Bischofia jacanica)、格木(Erythrophloeum fordii)、人面子(Dracontomelon duperreanum)、红椎(Castanopsis hystrix)、扁桃(Magifera persiciformis)、深山含笑(Michelia maudiae),同种植物苗龄、高度、基径和长势基本一致,并出自同一苗圃。
2) 处理方法:2006年3月从同一苗圃地选择相应实验树种的带土袋苗,用相同的基质土壤进行盆栽,口径为25 cm,内装土层厚度25 cm。经过38天相同环境(土壤基质、光照、水分管理)适应后,于2006年5月18—24日分别放置到每个试验点野外开阔区的坚实地面或地面铺1层塑料薄膜,每个点每个植物种5个重复,水分管理措施一致(用当地的地下水浇灌)。经过(830±6)天的暴露后,于2008年8月26—29日进行土壤样的采集。
3) 采集每个地点每个树种的根际土和非根际土土样采用Riley等(1971)的方法来区分根际土和非根际土。
1.2 长期在不同污染胁迫下的土壤样品采集本研究选取南亚热带区分布最广、最具代表性的树种(荷木)进行分析。分别在酸污染严重区的黄埔石化厂和相对清洁区的流溪河挖取荷木(10~15年)的根际及非根际土壤样品,同一地方同一树种分别选3株大小相同(或相近)的树,每株树在3个点采集根际和非根际土壤,则共9个根际土和9个非根际土采样点,然后将同一地点、同一土类(根际和非根际土)的土样混合成1个;进行样品处理和分析。两地的成土母岩都为花岗岩,土壤类型都为赤红壤。
1.3 土壤样品的化学测定方法1) 土壤pH值采用电位法(土水比为1:2.5)测定(国家林业行业标准LY/T 1239-199,下同)。2)土壤有机质采用重铬酸钾氧化-外加热法(LY/T1237-199)测定。3)土壤交换性钙、镁的测定采用1 mol·L-1乙酸铵交换-ICP测定,即土壤样品通过1 mol·L-1乙酸铵溶液处理后的浸出液(LY/T 1243-199),用ICP-OES(Varian Vista-MPX)测定,标准溶液中应加入同量的1 mol·L-1乙酸铵溶液,以消除基体效应。4)土壤交换性钾和钠的测定采用乙酸铵交换-火焰光度法(LY/T 1243-199)。5)土壤交换性酸、交换性Al3+的测定采用1 mol·L-1氯化钾交换-中和滴定法(LY/T 1240-199)。6)土壤Cl-,NO3-的测定按土水比1:5的比例提取后,采用离子色谱测定(ICS-90)。7)土壤硫酸根采用硫酸钡比浊法(LY/T 1251-199)测定。8)土壤铵态氮采用氧化镁浸提-扩散法(LY/T 1231-199)测定。
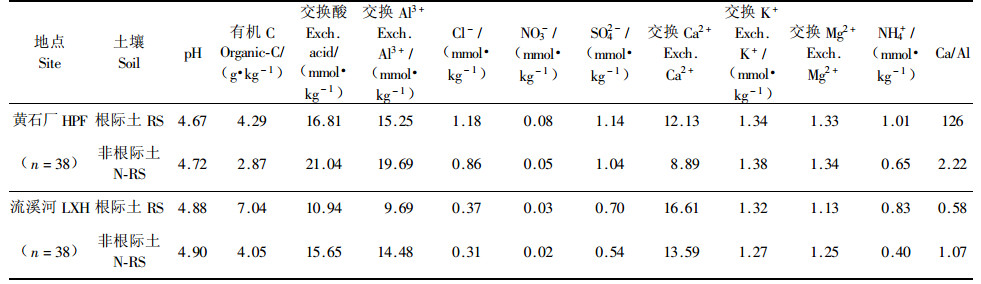
2 结果分析 2.1 植物短期暴露在污染环境下的土壤化学变化从图 1和表 1可知:1)pH值:总是根际土的pH值要比非根际土pH值小(即H+多),表现出根际酸环境特点;在胁迫不同的区域(酸污染区的黄埔石化厂试验点、相对清洁区的流溪河试验点),所有参试树种的根际土和非根际土中pH值总是污染区>相对清洁区,说明酸雨胁迫的提高引起土壤酸化的加强。2)SO42-等酸性物质:与H+相同,主要酸性阴离子(Cl-,NO3-,SO42-)总是根际土的含量要比非根际土的含量高;所有参试树种的根际土和非根际土中Cl-,NO3-,SO42-含量总是表现为酸雨污染区小于相对清洁区,这也是酸雨胁迫的具体表现。3)交换Al3+和交换酸:土壤交换Al3+和交换酸存在高度相关,不论在那种植物根际或非根际土壤、也不论在何种酸雨污染胁迫下,随着Al3+的增加,交换酸呈同步增加(图 2),且交换酸与Al3+之间关系达极显著水平(R2=0.996 3)。4)有效养分元素:从图 1和表 1可知:在酸雨污染胁迫区的黄埔石化厂,参试植物根际和非根际土壤中交换Ca2+含量(12.13和8.89 mmol1/2Ca2+·kg-1)明显低于酸污染胁迫小的流溪河相应植物根际和非根际土壤中交换Ca2+含量(16.61和13.59 mmol1/2Ca2+·kg-1),说明酸雨胁迫增加Ca2+的淋失,但在酸雨重胁迫环境下植物对Ca2+的吸收效率提高了,Ca2+从非根际土转移到根际土的速率或量增加(酸雨污染重胁区根际土Ca2+含量是非根际土含量的1.36陪,而轻胁区根际土Ca2+含量是非根际土含量的1.22陪);与Ca2+相似的还有有机碳。在污染胁迫区的黄埔石化厂,参试植物根际和非根际土壤中交换酸含量(16.81和21.04 mmol·kg-1)、NH4+含量(1.01和0.65 mmol·kg-1)明显高于酸污染胁迫小的流溪河对应交换酸含量(10.94和15.65 mmol·kg-1)和NH4+含量(0.83和0.40 mmol·kg-1),在酸雨胁迫增强时,植物根际和非根际土壤中NH4+和交换酸含量明显增加。5)元素间关系:76个短期暴露试验的植物根际和非根际土壤样品的交换Ca2+和Al3+成相对紧密的负相关趋势(R2=0.6606), 有机质和Al3+也成较好的负相关(R2=0.552 5)。

|
图 1 酸污染区和相对清洁区参试树种的根际土和非根际土主要成分化学含量 Figure 1 Chemical contents of rhizosphere and non-rhizosphere soil of experimental tree species at acid polluted site (HPF) and relative clean site (LXH) ◆黄埔石化厂试验点根际土Rhizosphere soil at Huangpu petrifaction factory (HPF); □流溪河试验点根际土Rhizosphere soil at Liuxihe(LXH); ▲黄埔石化厂试验点非根际土Non-rhizosphere soil in HS at HPF; ×流溪河试验点非根际土Non-rhizosphere soil in LXH |
|
|

|
图 2 植物暴露试验全部根际和非根际土壤中交换酸和交换Al3+之间关系 Figure 2 Relationship between exchangeable acid and exchangeable Al3+ for all rhizosphere and non-rhizosphere soil samples of the plant exposure experiment at acid polluted site (HPF) and relative clean site (LXH) |
本暴露试验结果说明:植物为短期暴露在酸雨污染环境下,土壤的重要化学响应:除植物体(根尖)增加各种有机酸分泌物的排放来与根区Al3+络合外,同时还通过增加根际土壤的有机质和NH4+含量、提高对交换性Ca2+的吸收效率等方式来削弱环境中不断增加的酸胁迫(SO42-,NO3-,Cl-和有机酸),并排斥Al3+在根际土壤环境的积累或减弱Al3+的毒性。
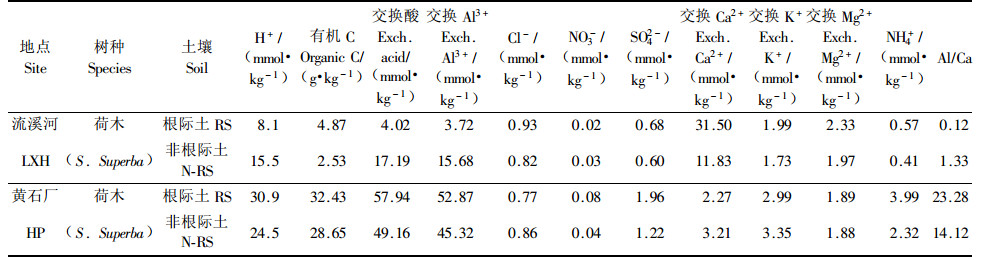
2.2 长期在不同污染胁迫下的土壤化学响应在长期酸雨胁迫下,荷木的根际和非根际土壤化学表现与植物暴露实验比较相似的结果。(表 2) 1)H+:相对清洁区的流溪河试验点荷木根际和非根际土取自近原始的天然林,根际土pH值(5.09)略微比非根际土(4.81)高,而在酸雨污染胁迫下(黄石厂)的荷木,总是根际土的H+要比非根际土H+多(即pH值小),表现出根际酸环境特点;不论是根际土还是非根际土,污染区的H+要比相对清洁区大,说明酸雨胁迫的提高引起土壤酸化的加强。2)SO42-等酸性物质:主要酸性阴离子(Cl-,NO3-,SO42-)总是根际土的含量要比非根际土的含量高;并且在污染区的含量大于在相对清洁区的含量。3)交换Al3+和交换酸:交换Al3+和交换酸存在唇齿相依关系,污染区的Al3+明显比相对清洁区高,随着Al3+的增加,交换酸呈同步增加,这与短期暴露试验的结果完全一致。不同的是在长期严重酸雨胁迫下(黄石厂),荷木的根际富集了大量的Al3+,使得根际土Al3+含量(52.87 mmol1/3Al3+·kg-1)比非根际土壤的Al3+含量(45.32 mmol1/3Al3+·kg-1)高,这也证实并支持了Samuels等(1997)得出的“Al很容易在植物根尖富集并抑制根的生长”的研究结论(Samuels et al, 1997),由于Al3+含量的增加,使得植物分泌更多的有机酸与根区Al3+络合,降低铝的毒性;而在相对清洁区(流溪河),由于没有受到更大酸雨污染胁迫,植物根际和非根际土中有机酸含量、Al3+含量仍然与短期暴露试验的结果一致,表现为根际土小于非根际土。4)有效养分元素:与短期暴露试验的结果完全一致是,酸污染高胁迫区(黄石厂)相比酸雨污染低胁迫区(流溪河),根际和非根际土壤交换Ca2+,Mg+有效养分明显下降,而NH4+、有机质、交换K明显增加。

|
图 3 植物暴露试验全部根际和非根际土壤中交换Ca2+和交换Al3+(A)、有机质与交换Al3+(B)之间关系 Figure 3 Relationship between exchangrable Ca2+ and exchangeable Al3+ (A), organic matter and exchangeable Al3+ (B) for all rhizosphere and non-rhizosphere soil samples of the plant exposure experiment at acid polluted site (HPF) and relative clean site (LXH) |
|
|
酸雨使土壤的pH值降低、营养阳离子析出、营养阴离子增加、营养物循环和供给发生变化,持续酸雨淋溶导致植物可利用营养元素减少,这些对植物的营养代谢和其他生理代谢产生影响(冯宗炜, 1993; 1999,Abrahamsen et al., 1994; Hagedorn et al., 2001; 樊后保等, 2002; 田大伦等, 2007),认为,土壤中的Ca2+和Mg2+等的淋失量随酸雨酸度增加而增加;但土壤对酸雨污染的响应明显地受土壤物理—化学性质的制约(Delfosse et al., 2004)。
本研究认为:土壤中主要离子(H+,Al3+,Cl-,NO3-,SO42-,K+,NH4+)含量在酸雨严重胁迫区(黄石厂)要大于相对清洁区(流溪河)的含量,这与Majdi等(1995)的试验结果中增加N和S的供应使根际和非根际土壤交换性元素含量提高结果一致;同时发现酸雨胁迫的加强使得有效养分尤其Ca2+的淋失增加,但也可以看出在酸雨重胁迫环境下植物对Ca2+的吸收效率提高,Ca2+从非根际土转移到根际土的速率或量增加;从根际和非根际土壤养分比较可知,根际土含量高于非根际土含量,这与荒漠环境胁迫下灌木的根际效应相似(詹媛媛等, 2009),但有些试验结果表现出根际土的K被植物吸收消耗使得含量比非根际土小(Zhang et al., 2002),这可能与植物对K大量吸收有关。
3.2 土壤化学物质间的拮抗作用本研究指出,根际和非根际土壤的交换Ca2+和Al3+存在拮抗作用即成相对紧密的负相关趋势(R2=0.660 6), 有机质和Al3+也存在诘抗作用(R2=0.552 5),说明有机质、Ca2+与Al3+关系紧密,在土壤对酸雨污染的缓冲中起最主要作用,这与岑慧贤等(2000)研究结果相同;同时也有研究指出对有些土壤Mg2+在酸雨污染的缓冲中起着最主要作用(Helina, 1996)。因此,如Huang等(1996)的研究结果,在Al对植物影响过程中,Al与Ca2+,Mg2+等离子有竞争与拮抗作用,Al对植物的伤害取决于各离子量的关系。
3.3 土壤铝毒分析酸雨对森林影响的铝毒假说最早是以Ulrich(1980)为代表的学者们所提出,并认为当土壤中Al/Ca值大于5 (钙/铝比率低于0.15)时,所溶出的铝具有毒性,将对植物产生危害(樊后保,2003),影响植物的养分营养代谢。在铝对植物毒性及抗性机制等方面的研究中,如Samuels等(1997)研究表明:Al很容易在植物根尖富集,抑制根的生长;Huang等(1992)研究指出Al抑制细胞膜对阳离子的运输;Minocha等(1992)研究指出Al能抑制DNA合成有关的酶活性。铝在土壤中以固定态存在,当土壤盐基饱和度下降到一定程度或土壤pH降到铝缓冲范围(pH2.8~4.2)时部分外源质子使矿物铝活化而释放Al3+(赵忠,1988;刘厚田等,1992);土壤在pH<3.0以后硅铝释放量骤增(董汉英,2000)。
本研究发现:在植物短期暴露实验中,严重酸雨污染区的根际和非根际土壤Al/Ca比值(交换Al和Ca交换的比值,下同)比相对清洁区大(表 1),说明酸雨明显引起Al的释放;但由于植物在污染胁迫下的暴露时间短(830±6)天,所以土壤Al/Ca比值小于5,未产生铝毒。而在长期污染胁迫下,酸雨污染低胁迫区(流溪河)的土壤Al/Ca比值小于5,严重酸雨胁迫下(黄石厂)的土壤Al/Ca比值小于5(表 2)。同时发现,无论在植物短期暴露实验或者植物在长期酸雨污染下,根际和非根际土壤的Al3+与有机酸存在非常紧密关系,因此本研究结果同意并支持Cambraia等(1983)、Pellet等(1996)研究观点:有机酸是去除Al毒性的重要途径,它与根区Al3+络合,降低铝的毒性。另外,在酸污染胁迫小时,根际土的Al3+和有机酸含量比在非根际土中的含量小,而当酸污染胁迫增强时则表现为根际土的Al3+和有机酸含量要比非根际土的大,且在在程度胁迫时发生转变,尚需要进一步研究。
岑慧贤, 仇荣亮, 杨平. 2000. 土壤酸沉降缓冲机制的探讨[J]. 环境科学研究, 13(2): 49-54. DOI:10.3321/j.issn:1001-6929.2000.02.014 |
岑慧贤, 王树功, 仇荣亮, 等. 2001. 模拟酸雨对土壤盐基离子的淋溶释放影响[J]. 环境污染与防治, 23(1): 13-15, 26. DOI:10.3969/j.issn.1001-3865.2001.01.005 |
董汉英, 仇荣亮, 吕越娜. 2000. 模拟酸雨对南方土壤硅铝释放的影响[J]. 环境科学, 21(1): 75-77. |
樊后保, 林德喜. 2002. 模拟酸雨对福建四种山地土壤的淋溶与风化作用[J]. 山地学报, 20(5): 570-577. DOI:10.3969/j.issn.1008-2786.2002.05.011 |
樊后保. 2003. 酸雨与森林衰退关系研究综述[J]. 福建林学院学报, 23(1): 88-92. DOI:10.3969/j.issn.1001-389X.2003.01.021 |
冯宗炜. 1993. 酸雨对生态系统的影响——西南地区酸雨研究[M]. 北京: 中国科学技术出版社.
|
冯宗炜. 1999. 酸沉降对生态系统的影响及其生态恢复[M]. 北京: 中国环境科学出版社.
|
高太忠, 戚鹏, 张扬, 等. 2004. 酸雨对土壤营养元素迁移转化的影响[J]. 生态环境, 13(1): 23-26. DOI:10.3969/j.issn.1674-5906.2004.01.008 |
刘厚田, 田仁生. 1992. 重庆南山马尾松衰亡与土壤铝活化的关系[J]. 环境科学学报, 12(3): 297-304. DOI:10.3321/j.issn:0253-2468.1992.03.008 |
罗微, 林钊沐, 茶正早, 等. 2004. 热带作物磷肥根际生物有效性研究进展[J]. 热带农业科学, 24(5): 85-91. DOI:10.3969/j.issn.1009-2196.2004.05.018 |
田大伦, 黄智勇, 付晓萍. 2007. 模拟酸雨对盆栽樟树(Cinnamomum camphora)幼苗叶矿质元素含量的影响[J]. 生态学报, 27(3): 1099-1105. DOI:10.3321/j.issn:1000-0933.2007.03.034 |
詹媛媛, 薛梓瑜, 任伟, 等. 2009. 干旱荒漠区不同灌木根际与非根际土壤氮素的含量特征[J]. 生态学报, 29(1): 59-66. DOI:10.3321/j.issn:1000-0933.2009.01.008 |
赵忠. 1988. 森林土壤酸化及其对林木生长的影响[J]. 土壤学进展, (2): 1-5. |
Abrahamsen G, Stuanes A O, Tveite B. 1994. Long-term experiments with acid rain in Norwegian forest ecosystem[M]. Berlin: Springer-Verlag.
|
Cambraia J, Galvan F R, Estevao M M. 1983. Effects of aluminum on organic acid, sugarandamino acid composition of the root system of sogghum[J]. Journal of Plant Nutrition, 6: 313-322. DOI:10.1080/01904168309363092 |
Delfosse T, Delmelle P, Iserentant A, et al. 2004. Contribution of SO3 to the acid neutralizing capacity of Andosols exposed to strong volcanogenic acid and SO2 deposition[J]. European Journal of Soil Science, 55: 1-13. DOI:10.1046/j.1365-2389.2004.00590.x |
Hagedorn F, Maurer S, Egli P, et al. 2001. Carbon sequestration in forest soils: effects of soil type, atmospheric CO2 enrichment, and N deposition[J]. European Journal of Soil Science, 52: 619-628. DOI:10.1046/j.1365-2389.2001.00412.x |
Helina H. 1996. Soil response to acid percolation[J]. J Environ Qual, 25(4): 638-645. |
Huang J W, Pellet D M, Papernik L A. 1996. Aluminium interactions with voltage dependant calcium transport in plasmamembrane vesicle isolated from roots of aluminum sensitive and resistant wheat cultivars[J]. Plant Physiology, 110: 561-569. DOI:10.1104/pp.110.2.561 |
Majdi H, Bergholm J. 1995. Effects of Enhanced Supplies of Nitrogen and Sulphur on Rhizosphere and Soil Chemistry in a Norway Spruce Stand in SW Sweden[J]. Water, Air and Soil Pollution, 85: 1777-1782. DOI:10.1007/BF00477237 |
Minocha R, Minocha S C, Long S L, et al. 1992. Effect of aluminum on DNA synthesis, Cellular polyamines, polyamine biosynthetic enzymes and inorganic ions in cell suspension cultures of a wood plant, Catharanthus roseus[J]. Physiologia Plantanum, 85: 417-424. DOI:10.1111/ppl.1992.85.issue-3 |
Pellet D M, Papernik L A, Kochian L V. 1996. Multiple aluminum resistance mechan isms in wheat, Roles of rootapical phose phate and malate exudation[J]. Plant Physiology, 112: 591-597. DOI:10.1104/pp.112.2.591 |
Riley D, Barber S A. 1971. Effect of ammonium and nitrate fertilization on phosphorus uptake as related to root induced pH changes at the root-soil interface[J]. Soil Sci Soc Am Proc, 35: 301-306. DOI:10.2136/sssaj1971.03615995003500020035x |
Samuels T D, Kucukakyuz K, Magaly R Z. 1997. Al partitioning patterns and root growth as related to Al sensitivity and Al tolerance in wheat[J]. Plant Physiology, 113: 527-534. DOI:10.1104/pp.113.2.527 |
Zhang Z L, Eckhard G. 2002. Changes in the extractability of cations (Ca, Mg and K) in the rhizosphere soil of Norway spruce (Picea abies) roots[J]. Plant and Soil, 243: 209-217. DOI:10.1023/A:1019915512072 |