 2009, Vol. 45
2009, Vol. 45文章信息
- 李庆梅, 陈怀梁, 马风云, 刘艳, 张俊风.
- Li Qingmei, Chen Huailiang, Ma Fengyun, Liu Yan, Zhang Junfeng
- 云杉种子超干贮藏的生理生化特征
- Effects of Ultra-Dry Treatment on Physiological and Biochemical Characteristics of Picea asperata Seeds
- 林业科学, 2009, 45(11): 50-54.
- Scientia Silvae Sinicae, 2009, 45(11): 50-54.
-
文章历史
- 收稿日期:2009-07-10
-
作者相关文章


2. 山东农业大学林学院 泰安 271018;
3. 北京林业大学森林培育与保护教育部重点试验室 北京 100083
2. Forestry College of Shandong Agricultural University Tai'an 271018;
3. Key Laboratory for Silviculture and Conservation of Ministry of Education Beijing Forestry University Beijing 100083
超干贮藏以其低廉的造价越来越受到研究者的重视,种子贮藏时最佳含水量的确定及超干后种子生理生化特征成为当今种子界研究的热点(Zheng et al., 1998;李庆梅等,2009)。超干种子耐贮性机理研究的目的,就是想弄清楚种子质变的机理,尽可能简化贮藏条件,降低贮藏成本,使种子在简单处理后能存放得更久(Ellis et al., 1990)。1985年,国际植物遗传资源委员会首先提出对某些作物种子采用超干贮藏的设想。Ellis等(1989)将芝麻(Sesamum indicum)种子含水量由50 g·kg-1降到20 g·kg-1,种子寿命延长了40倍。20世纪80年代后期,国内一些农业院校、研究所相继开展了超干种子研究并取得了一些研究成果:刘岩等(1990)首先在我国开展超干种子贮藏研究,在超干种子的生理和细胞学方面取得了一定进展;其他研究者结合超干种子抗老化能力,对种子的脱氢酶、超氧歧化酶、过氧化物酶、淀粉酶、自由基等生理生化特性进行了研究(朱诚等,2001;郑郁善等,2002;李毅,2007),并对超干种子是否采取回湿措施进行了比较,发现了超干种子耐贮藏性与生理生化指标的相关性(张云兰等,1994)。
云杉(Picea asperata),松科(Pinaceae)云杉属常绿乔木,分布于云南、四川、甘肃等地,是西南高山区主要造林树种之一。结实时间较长(30~40年),结果盛期出现在60~120年,种子年大致每3~5年出现1次,种子贮藏年限较短(自然条件下保存3年)。近年来种子丰年间隔越来越长,球果锈病在中国局部地区的蔓延也严重影响云杉种子收成,所以云杉种子产量越来越低,生产上出现了供不应求的现象。本研究首次研究了超干燥及老化处理对云杉种子活力和生理生化特性的影响,探讨云杉种子超干贮藏的特性及生理生化变化规律,为保证云杉造林用种子的永续利用提供理论基础,也为其他林木种子的超干贮藏提供参考。
1 材料与方法 1.1 试验材料云杉种子由中国林木种子公司提供,产地云南,采期为2007年秋。初始含水量为82 g·kg-1,千粒质量为5.04 g,发芽率为83.3%。
1.2 试验方法 1.2.1 超干种子制备1) 硅胶室温干燥取适量种子用纱布袋包好,放在下层为硅胶的密封干燥器中。硅胶与种子质量比为10:1,每天更换经120 ℃烘干冷却至室温的干硅胶。每天定时称质量,制备不同含水量的种子。将不同含水量的种子分别密封于密闭容器中(郑郁善等,2002;Hou et al., 2008)。2)种子回湿处理测定指标前,先将干燥处理的种子逐级回湿,置于底部盛有饱和CaCl2水溶液的干燥器中(相对湿度35%),在室温下回湿处理2天,然后转移至盛有饱和NaCl水溶液的干燥器中(相对湿度75%)处理2天,取出于室温条件保存(李庆梅等,2009)。3)种子老化处理系列含水量种子回湿后放入密闭容器中,置于50 ℃生化培养箱中(RH为100%)老化7天。
1.2.2 种子发芽率和活力测定按GB2772-1999《林木种子检验规程》(国家质量技术监督局,1999)的技术标准进行。3个重复,每个重复100粒种子,种子经45 ℃温水自然冷却浸泡24 h,用去离子水充分冲洗干净,放在铺有2层滤纸的培养皿中置于25 ℃培养箱进行发芽试验,每天补充水分并记录发芽种子数,在第10天统计发芽势,第24天结束发芽试验,统计发芽率。发芽指数(GI)按如下公式计算:GI=∑(Gt/Dt),式中Gt为t日后的发芽数,Dt为发芽日数。
1.2.3 电导率的测定取100粒种子(大小均匀饱满且无损伤),用去离子水冲洗数次,吸干表面水分,置带盖的器皿中,加100 mL去离子水,于30 ℃恒温水浴中浸泡2,4,6,8,10 h,用电导仪(delta326)测定浸泡液电导率(Ci,i=2,4,6,8,10),然后加热煮沸15 min,冷却至室温测定电导率(C0)。重复3次。相对电导率按照以下公式(宋松泉等,2005)计算:相对电导率=Ci/C0×100%。
1.2.4 脱氢酶活性测定每个样品取云杉种子100粒称质量,3个重复,45 ℃温水室内自然冷却浸种24 h(室温20~25 ℃),10 mL 1 g·kg-1 TTC溶液(现用现配)35 ℃的恒温(加盖保持黑暗条件)反应10 h,取出研磨,丙酮定容至10 mL,4 000 r·min-1下离心10 min,上清液用TU-1801紫外可见分光光度计在波长490 nm处测定密度值,根据标准曲线得出还原态TTC值(李庆梅等,2009)。
1.2.5 异柠檬酸裂解酶活性测定取50粒种子经45 ℃温水自然冷却浸泡24 h后,在25 ℃下萌发3天,置研钵中,加入3 mL预冷的Tris-HCl缓冲液,在水浴中研磨成糊状,3000r·min-1离心5 min,吸取上清液,并用1 mL缓冲液冲洗沉淀,3000 r·min-1再离心5 min,吸取上清液,弃沉淀,合并上清液。12000 r·min-1离心15 min,上清液即为酶液,将上清液定容至10 mL,立即测定异柠檬酸裂解酶(ICL)活性。取2支试管,分别加入1 mL缓冲液,0.2 mL 150 μmol·L-1硫酸镁,0.2 mL 60 μmol·L-1半胱氨酸盐酸,0.2 mL酶液。30 ℃恒温水浴中保温10 min,再加入0.2 mL 7 mmol·L-1异柠檬酸三钠,30 ℃恒温水浴中反应30 min,加入2 mL 1.5 mol·L-1 NaOH显色,另以1 mL缓冲液,加0.2 mL 0.1%二硝基苯肼和2 mL 1.5 mol·L-1 NaOH作空白对照,在波长445 nm下比色,读取光密度值。在标准曲线上还原乙醛酸含量(张北壮,1988)。
1.3 数据分析数据方差分析和多重比较采用SPSS11.5辅助完成,图应用Microsoft Excel辅助完成。
2 结果与分析 2.1 云杉种子干燥处理发芽生理指标的变化及回湿后抗老化能力由表 1看出,云杉种子含水量由82 g·kg-1降至64,52,41,29 g·kg-1再经回湿处理后,其发芽率和活力均有显著的变化。种子含水量在82~41 g·kg-1范围时,种子的发芽势、发芽率和发芽指数均维持在较高的水平,当含水量降至29 g·kg-1时,种子发芽率骤然降至60%左右,其发芽势、发芽率和发芽指数均显著低于其他含水量种子。
|
|
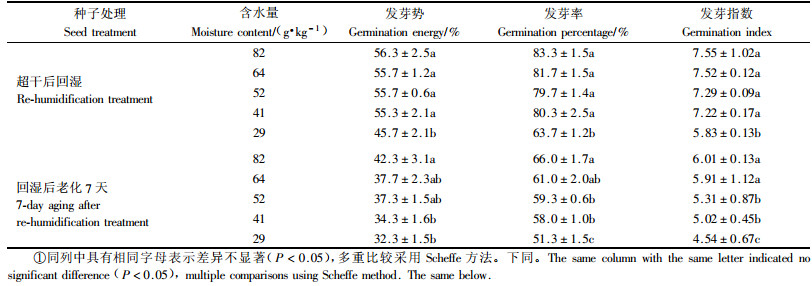
超干回湿后的种子老化(50 ℃,RH为100%)7天后,不同含水量种子的发芽势、发芽率和发芽指数指标均比老化前低,含水量64,52和41 g·kg-1的各项指标与82 g·kg-1差异显著,含水量29 g·kg-1与82 g·kg-1差异极显著(表 1)。可见自然含水量种子的抗老化能力最高,含水量降低后种子发芽率与活力有所降低,含水量64,52,41 g·kg-1的云杉种子还具有一定的抗老化能力,而29 g·kg-1的含水量完全不能接受。
2.2 超干处理对种子细胞膜结构的影响种子发生劣变时,细胞膜结构和功能的完整性受到伤害,导致膜透性增加。因此种子会向周围的溶液中渗漏出较多的电解质。近年来一些种子保存特性的研究也采用相对电导率(Li et al., 2007),一方面避免了所选种子自身差异造成对观测效应的混淆,另一方面能够直观地描述细胞膜由半透膜变为通透膜时透出电解质的差异(李庆梅等,2009)。当云杉种子含水量由82 g·kg-1降至29 g·kg-1再经回湿处理后,其相对电导率有显著的变化(图 1)。超干种子含水量在82~41 g·kg-1范围时,种子的相对电导率保持在一较低水平上,当含水量降至29g·kg-1时,相对电导率要显著高于其他含水量种子,其细胞膜的选择透性最差,超干后的回湿措施没有对细胞膜产生显著修复,这说明回湿措施对其细胞膜的修复效果不佳。

|
图 1 云杉种子老化前后相对电导率 Figure 1 The relative conductivity of seeds pre-aging and after 7 day aging |
云杉种子回湿老化7天后的相对电导率较老化前高,说明种子细胞膜的选择透性不同程度地受到损伤,总的趋势是种子的相对电导率随着含水量的降低而升高,种子细胞膜损伤程度加重,自然含水量种子在老化后相对电导率最低,说明在老化过程中其细胞膜的选择性受到的损伤最小。
2.3 老化处理对云杉超干种子脱氢酶活性的影响种子脱氢酶活性与种子活力关系密切,是种子发芽率和活力的最重要生理指标之一,可反映细胞代谢的还原能力和胚损伤程度(李庆梅等,2009)。试验结果(图 2)表明,种子含水量在82~41 g·kg-1范围时,其脱氢酶活性无显著差异,基本保持在同一个水平。当含水量降至29 g·kg-1时种子脱氢酶活性降幅较大,种子活力下降较大,这与发芽试验的结果相吻合(表 1)。

|
图 2 云杉种子老化前后脱氢酶活性 Figure 2 The dehydrogenase activity of seeds pre-aging and after 7-day aging |
回湿后老化7天,含水量不同的云杉种子脱氢酶活性差异显著,均值比较,自然含水量种子酶活性最高,与含水量52,41,29 g·kg-1差异显著,而且随含水量的降低酶活性也呈下降趋势,含水量为29 g·kg-1的超干种子酶活性最小。
2.4 老化处理对种子异柠檬酸裂解酶活性的影响油类种子在萌发过程中,通过脂肪酸的β-氧化和乙醛酸循环将脂肪酸转变成碳水化合物。在乙醛酸循环中,异柠檬酸裂解酶(ICL)是关键酶之一,它催化异柠檬酸裂解成乙醛酸和琥珀酸,前者进入乙醛酸循环,后者通过一系列反应主要是糖酵解的逆转合成葡萄糖。因此,测定ICL活性对了解油类种子的代谢途径和物质转化,以及种子活力情况有重要意义。当云杉种子含水量由82 g·kg-1降至29 g·kg-1再经回湿处理后,其ICL活性有显著的变化(图 3)。种子含水量在82~41 g·kg-1范围时,种子的ICL活性有先下降后上升的规律,但酶活性之间差异不显著,当含水量降至29 g·kg-1时ICL活性要显著低于其他含水量种子。

|
图 3 云杉种子老化前后ICL活性 Figure 3 The ICL activity of seeds pre-aging and after 7-day aging |
回湿后老化7天,含水量不同的云杉种子ICL活性差异显著,均值比较,自然含水量种子ICL活性最高,与含水量52,41,29 g·kg-1差异显著,而且随含水量的降低酶活性也呈下降趋势,含水量为29 g·kg-1的超干种子酶活性最小。
3 结论与讨论 3.1 超干对种子发芽指标的影响云杉含水量在82~41 g·kg-1范围时,种子的发芽率、发芽势和发芽指数都保持在较高水平,含水量降至29 g·kg-1,其发芽率、发芽势和发芽指数都有大幅下降,与其他含水量的发芽指标有显著差异,说明该种子适宜在82~41 g·kg-1范围内保存。李庆梅等(2009)研究发现当油松(Pinus tabulaeformis)含水量降至32 g·kg-1时,仍保持较高的发芽活力;郑郁善等(2002)研究发现马尾松(Pinus massoniana)含水量50g·kg-1时发芽率和各项活力指标达到最大,黑松(Pinus thunbergii)含水量40 g·kg-1时发芽率和各项活力指标达到最大;侧柏(Platycladus orientalis)为37 g·kg-1(Hou et al., 2008);杜仲(Eucommia ulmoides)为35 g·kg-1(林坚等,1996);也有研究者指出种源地和种质对林木种子最低保存含水量也有影响(张云兰等,1994;Hou et al., 2008)。
朱诚等(2001)对超干洋葱(Allium fistulosum)种子的研究指出,种子贮藏的最适含水量随温度的改变而发生相应的变化,35 ℃最适含水量为34g·kg-1,室温下为34~35g·kg-1,15 ℃为45~51g·kg-1。多数林木种子都有坚硬的种壳,对胚和胚乳具有较好的保护作用,林木种子贮藏的最适含水量是否随温度的改变而发生相应的变化,有待于进一步研究。
3.2 超干对膜系统的影响当云杉种子含水量由82 g·kg-1降至29 g·kg-1再经回湿处理后,其相对电导率有显著的变化(图 1)。超干种子含水量在82~41 g·kg-1范围时,种子相对电导率保持在较低水平,当含水量降至29g·kg-1时,相对电导率显著高于其他含水量种子,细胞膜选择透性最差。种子含水量降低,细胞膜中磷脂的排列发生变化,电镜观察结果表明,膜结构的变化包括膜磷脂分子由双层排列状态变成六角形构相,细胞膜从流动相变成凝胶相,从而破环了细胞膜的渗透功能。当回湿时,膜结构又恢复原状,其中膜构相恢复过程所需的时间以及最终整合的完善程度,决定着种子的活力水平(程红焱等,2005)。当含水量降至29 g·kg-1时,相对电导率最高,这说明回湿措施对其细胞膜的修复效果不佳。
3.3 超干对酶活性的影响脱氢酶是对种子染色的定量测定,脱氢酶活性与种子活力之间的关系很密切,可反映细胞代谢的还原能力和胚损伤程度。云杉种子干燥至41 g·kg-1,种子脱氢酶活性保持稳定状态,说明种子含水量降至50 g·kg-1以下,对种子脱氢酶活性没有产生显著性影响。脱氢酶活性随种子衰老而降低,在种子生理代谢过程中脱氢酶受老化影响其活性下降,几乎与种子生活力的下降相平衡。程红焱等(1992)在测定榆树(Ulmus pumila)种子脱氢酶活性时,观测到伴随着种子的衰老,脱氢酶活性显著降低,并且下降的速率与发芽力的降低相一致。对马尾松(郑郁善等,2001)、杉木(Cunninghamia lanceolata)(郑郁善等,2002)、银合欢(Leucaena leucocephala)(邹冬梅,2006)、侧柏(Hou et al., 2008)、油松(李庆梅等,2009)等研究均获得相似的结果。
油类种子萌发时,通过脂肪酸的β-氧化和乙醛酸循环将脂肪酸转变成碳水化合物。在乙醛酸循环中,异柠檬酸裂解酶(ICL)是关键酶之一。异柠檬酸裂解酶催化异柠檬酸的分解,形成1分子乙醛酸和1分子琥珀酸。前者和二硝基苯肼作用形成乙醛酸苯肼,在碱性条件下显褐色,其颜色深浅与乙醛酸含量呈正比。因此可以用比色法测定乙醛酸含量,并计算得出异柠檬酸裂解酶活性。超干种子含水量在82~41 g·kg-1时,种子异柠檬酸裂解酶活性之间差异不显著,当含水量降低至29 g·kg-1时,异柠檬酸裂解酶活性显著下降。本试验中异柠檬酸裂解酶活性与种子的发芽势的变化趋势保持一致,与胡家恕等(1994)对大豆(Clycine max)种子萌发的研究结论一致,即种子发芽势与种胚中异柠檬酸裂解酶活性呈极显著的线性正相关。
3.4 超干种子回湿后抗老化能力回湿后的超干种子在老化过程中发芽率、发芽势、发芽指数、相对电导率、脱氢酶活性及异柠檬酸裂解酶活性各项指标均低于自然含水量种子。回湿使得种子内部进行膜的修复活动,减慢种子劣变,提高种子活力,回湿后的种子在各项指标的测定过程中均表现出与自然含水量种子无明显差异。本试验采用回湿以后再进行高温高湿老化,回湿后,超干种子含水量均略高于自然含水量种子,所以回湿后的超干种子在老化过程中的衰变速率要大于自然含水量种子。张云兰等(1994)指出不同的种子应采用不同的回湿方法,回湿方法对种子发芽率和活力影响很大,绿豆(Phasedus aureus)用30%PEG处理16 h好于饱和水蒸汽回湿2天的效果,谷子(Setaria italica)以饱和水蒸汽回湿2天效果最好。
3.5 结论1) 云杉种子经过干燥处理后,种子含水量降至50 g·kg-1以下,从发芽率、发芽势、发芽指数、相对电导率、脱氢酶活性及异柠檬酸裂解酶活性综合分析表明,含水量在41g·kg-1以上时具有较好的耐脱水性,各项生理指标下降较小;含水量过低(29 g·kg-1),上述生理指标均明显低于对照和低含水量(41 g·kg-1)种子。超干种子的含水量为41 g·kg-1效果最好。2)回湿后种子含水量均略高于自然含水量种子,在老化试验中表现出耐贮性低于自然含水量种子,可能与回湿方法有一定的关系。3)试验未进行超干条件下的高温耐贮性比较。
程红焱, 郑光华, 景新明. 1992. 超干处理提高榆树种子的耐藏性(简报)[J]. 植物生理学通讯, 28(5): 340-342. |
程红焱, 宋松泉, 朱诚, 等. 2005. 超干种子耐贮藏性的细胞学及生理生化基础[J]. 云南植物研究, 27(2): 11-18. |
国家质量技术监督局.1999.林木种子检验规程.中华人民共和国国家标准GB 2772-1999.北京: 中国标准出版社. http://www.csres.com/detail/61893.html
|
胡家恕, 邵爱萍, 叶兆杰. 1994. 砷化镓和其它砷化物对大豆种子萌发和异柠檬酸(ICL)及超氧物歧化酶(SOD)活性的影响[J]. 农业环境保护, 13(5): 194-198. |
李庆梅, 侯龙鱼, 段新芳, 等. 2009. 油松种子超干贮藏生理生化特征的研究[J]. 林业科学研究, 22(1): 124-128. DOI:10.3321/j.issn:1001-1498.2009.01.022 |
李毅.2007.甘肃河西地区三种荒漠植物种质资源超干保存研究.兰州: 兰州大学博士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-10730-2007116064.htm
|
林坚, 郑光华, 程红焱. 1996. 超干贮藏杜仲种子的研究[J]. 植物学通报, 13(增刊): 58-62. |
刘岩, 陈杭, 郑光华. 1990. 种子吸湿-回干处理的应用及其原理[J]. 种子, (5): 34-35. |
宋松泉, 程红焱, 龙春林, 等. 2005. 种子生物学研究指南[M]. 北京: 科学出版社, 61-62.
|
张北壮. 1988. 种子中异柠檬酸裂解酶活性的测定[J]. 植物生理学通讯, (3): 68-70. |
张云兰, 郭新荣. 1994. 超干种子回湿方法研究[J]. 作物品种资源, (4): 48-50. |
郑郁善, 王舒凤, 陈礼光, 等. 2001. 马尾松等种子超干贮藏生理生化特性研究[J]. 林业科学, 37(1): 42-46. |
郑郁善, 王舒凤, 陈礼光, 等. 2002. 衬质处理提高超干贮藏林木种子活力研究[J]. 福建林学院学报, 22(2): 97-100. DOI:10.3969/j.issn.1001-389X.2002.02.001 |
朱诚, 曾广文, 胡家恕, 等. 2001. 超干洋葱种子抗老化作用及其自由基的清除[J]. 浙江大学学报:农业与生命科学版, 27(2): 139-144. |
邹冬梅. 2006. 银合欢种子的超干保存[J]. 热带作物学报, 27(2): 17-21. |
Ellis R H, Hong T D, Roberts E H. 1989. A comparison of the low-moisture-content limit to the logarithmic relation between seed moisture and longevity in twelve species[J]. Annals of Botany, 63(6): 601-611. |
Ellis R H, Hong T D, Roberts E H, et al. 1990. Low-moisture-content limits to relations between seed longevity and moisture[J]. Annals of Botany, 65(5): 493-504. DOI:10.1093/oxfordjournals.aob.a087961 |
Hou Longyu, Duan Xinfang, Li Qingmei, et al. 2008. Effects of ultra-drying on viability and physiological characteristics of Platycladus orientalis seed from arid and semiarid areas of northwest China[J]. Chinese Forestry Science and Technology, 7(1): 88-92. |
Li Yi, Hu Yuanfeng, Chen Tuo, et al. 2007. Physiological responses of Limonium aureum seed to ultra-drying[J]. Journal of Integrative Plant Biology, 49(5): 569-575. DOI:10.1111/jipb.2007.49.issue-5 |
Zheng Guanghua, Jing Xinming, Tao Karling. 1998. Ultradry seed storage cuts cost of gene bank[J]. Nature, 393: 223-224. DOI:10.1038/30383 |