 2009, Vol. 45
2009, Vol. 45文章信息
- 牟少华, 彭镇华, 孙启祥, 高志民, 李雪平.
- Mu Shaohua, Peng Zhenhua, Sun Qixiang, Gao Zhimin, Li Xueping
- 部分箬竹属植物的荧光AFLP分析
- Genetic Diversity of Indocalamus Determined by AFLP Marker
- 林业科学, 2009, 45(11): 32-35.
- Scientia Silvae Sinicae, 2009, 45(11): 32-35.
-
文章历史
- 收稿日期:2008-08-25
-
作者相关文章



2. 中国林业科学研究院林业研究所 北京 100091
2. Research Institute of Forestry, Chinese Academy of Forestry Beijing 100091
箬竹属(Indocalamus)是多年生禾本科(Gramineae)竹亚科(Bambusoideae)植物,其叶大、植株矮小、常绿、姿态优美,是理想的庭院观赏和园林绿化竹种。与其他竹类植物一样,箬竹属植物开花周期较长,形态分类主要是依靠营养体的特征,而营养体的特征往往会因为生长环境的影响,产生很大的变异(杨光耀等,2000),而且许多种的隶属关系还不清晰,种的划分也存在一些分歧(朱石麟等,1994;Ohrnberger,1999)。所有这些给该属植物的资源收集、种质鉴定、分类和引种带来了很大困难。
AFLP分子标记技术由荷兰科学家Zabeau等(1995)创立。该方法结合了RFLP和RAPD技术的特点,既具有PCR的高效性、安全性和方便性,又具有RFLP可靠性好、重复性高的优点,已被广泛应用于植物亲缘关系、种质鉴别及遗传多样性研究(王利等,2008;王献等,2005;Zhang et al.,2000;文亚峰等,2008;宋红竹等,2007)。
本研究采用AFLP技术分析16种箬竹以及形态相似的5个外源竹种,揭示其亲缘关系,并初步弄清了个别有争议种类的分类位置,为箬竹属植物的种质资源保存以及可持续利用提供科学依据。
1 材料与方法 1.1 材料每个竹种(表 1)经过专家鉴定后,在竹种的每个群体中随机选取20株,每株选取1片幼嫩的完全展开叶,取等量的组成混合样作为1个样品,用于试验分析。全部室内试验于国际竹藤网络中心国家林业局竹藤科学与技术重点开放实验室完成。
|
|
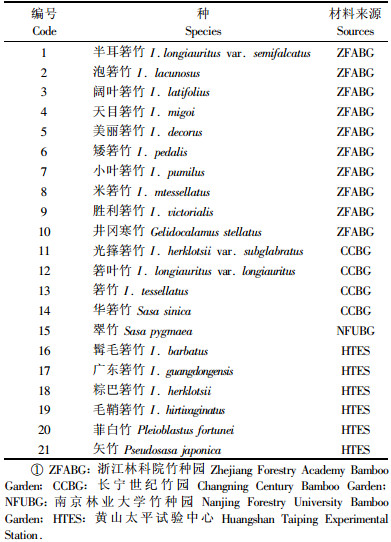
接头和引物由上海博亚生物技术有限公司(BIOASIA)北京分公司合成。Promega Tag DNA polymerase、Promega dNTP购于北京华绿渊生物技术公司;MseⅠ、EcoRⅠ和T4 DNA ligase为New England Biolabs公司产品。
1.2 方法 1.2.1 DNA提取总DNA提取采用CTAB方法(Hamilton et al., 1990)。分光光度计检测DNA浓度,稀释到50~100 ng·μL-1,-20℃保存备用。
1.2.2 AFLP凝胶电泳AFLP分析参照Vos等(1995)的方法,采用EcoRⅠ/MseⅠ酶切组合进行基因组限制性酶切(Barrett et al.,1998;田清震等,2000)。37℃酶切连接4 h以上。选择性扩增时DNA模板稀释20倍。AFLP预扩增反应选用EcoRⅠ00/MseⅠ00引物组合,选择性扩增反应采用(E+3)/(M+3)引物组合,其中E和M分别表示EcoRⅠ和MseⅠ引物的共同序列,E的共同序列为5′-GACTGCGTACCAATT-3′,M的共同序列为5′-GATGAGTCCTGAGTAA-3′。
选择扩增产物4%聚丙烯酰胺变性胶电泳,1 200 V电压电泳。电泳大约1 h,测序仪(ABI PRISM377 sequencer)激光管开始收集条带。电泳结束后,获得胶图。
1.2.3 数据分析用GENESCAN3.1软件开胶图,进行数据提取,安装胶图的MATRIX,选择合适的内标即SIZE STANDARD,分析得到结果。
通过Binthere软件设置片段大小的范围,导入数据,选择相应的荧光标记的颜色并选择合适的内标,导出结果并将结果保存为XLS格式。将表内的数值不为0的转换为1(数值为0的不转换),从而生成由“1”、“0”组成的原始矩阵。
采用NTSYSpc2.10e软件计算相似性系数与遗传距离系数,用SAHN Clustering进行不加权成对算术平均法(UPGMA)聚类分析。
2 结果与分析 2.1 AFLP指纹图谱从64对引物组合中筛选出9对扩增较整齐、多态性高的引物组合作为箬竹属植物选择性扩增的引物。这9对引物分别为:E-AAC/M-CAA,E-AGC/M-CAA,E-AAC/M-CAC,E-AAG/M-CAC,E-ACT/M-CAC,E-AAG/M-CAG,E-ACA/M-CTC,E-ACT/M-CTC和E-ACT/M-CTT。部分引物组合扩增结果见图 1。

9对AFLP引物在21个样品基因组中共获得1 367条清晰的谱带,其中共有条带115条,特异性带273条,特异性缺失条带72条。在16个箬竹样品基因组中共获得1 193条清晰的谱带,其中共有条带190条,特异性带177条,特异性缺失条带98条。这些特异性带或特异性缺失条带可作为识别不同竹种的分子标记。由此可见,在箬竹属植物的系统位置检测中,AFLP是非常有效的工具。
2.2 聚类结果与分析利用NTSYSpc 2.10e软件对AFLP数据进行分析,得到16种箬竹和5个形态相似竹种间的AFLP聚类图(图 2)。

|
图 2 箬竹属植物基于DICE相似系数的AFLP谱带UPGMA聚类图 Figure 2 The UPGMA dendrogram of Indocalamus using DICE coefficients |
采用DICE系数,当遗传相似性水平取阈值0.62时,可将样品分为2个大组。第1大组均为箬竹属植物,第2大组为华箬竹、矢竹、翠竹、菲白竹和井冈寒竹5种外源植物。第1大组的箬竹属植物又可以分为2组,一组为髯毛箬竹、广东箬竹、半耳箬竹、胜利箬竹、美丽箬竹、毛鞘箬竹、天目箬竹、阔叶箬竹、米箬竹、泡箬竹和矮箬竹,共11种箬竹;另一组为小叶箬竹、光箨箬竹、箬竹、箬叶竹和粽巴箬竹。其中,天目箬竹和阔叶箬竹相似系数较高,亲缘关系也较近,它们与毛鞘箬竹先聚在一起,然后再与米箬竹、泡箬竹聚成一小组。从图中还可以看出,半耳箬竹与箬叶竹、米箬竹与箬竹、光箨箬竹与粽巴箬竹关系都较远。第2大组中,翠竹与菲白竹相似系数较高,与华箬竹和矢竹聚成一小组,井冈寒竹与它们的关系较远。
3 结论与讨论 3.1 AFLP技术在箬竹属植物系统位置研究中的应用在供试的样品基因组中,9对AFLP引物分析结果表明,这些植物样品中都有共有的谱带、特异性带或特异性缺失条带,这些谱带可以作为识别箬竹种的分子标记。在箬竹属植物的系统学检测中,AFLP是一种非常有效的工具。
3.2 箬竹属植物的系统位置分析AFLP聚类结果表明,箬竹属16个种单独聚成一大组,而5个外源竹种聚成一组,这与形态分类上关于属的划分不相吻合。其中,菲白竹、翠竹和华箬竹与箬竹属植物营养体形态十分相似,通常认为赤竹属是高海拔竹种,箬竹属是低海拔竹种,但是2属的雄蕊数目、维管束形态不同(温太辉,1991)。从聚类结果看,两属的关系较远。
矢竹属的模式种矢竹每节仅一分枝,且与秆近等粗,这一特征与箬竹属十分接近,但它与箬竹属关系较远(杨光耀等,2000)。这得到本文结果的支持。
井冈寒竹具复轴型地下茎,竹秆灌木状,大型圆锥状真花序,顶生于无叶的侧生枝条顶端,雄蕊3,柱头2,这些特征与箬竹属很相似,有的学者曾将其归入箬竹属(Clayton et al., 1986)。但其竹秆每节分枝7~12,枝条纤细,不再分枝,顶端常仅具1叶,维管束为半开放型(温太辉,1991),出笋期为秋冬季等特征与箬竹属有异(耿伯介等,1996;王正平,1997)。本文分析结果也表明,井冈寒竹与箬竹属关系较远,不支持将其归入箬竹属。
在所有的箬竹属植物中,阔叶箬竹与天目箬竹相似性系数最高,研究结果支持耿氏分类系统(耿伯介等,1996),将天目箬竹归入阔叶箬竹中。同时,二者在形态上与毛鞘箬竹、米箬竹较相似,秆中部箨上的箨片基部均不向内收窄,箨耳不存在或稀可微弱发达。区别是毛鞘箬竹箨鞘的上部包秆甚紧贴,不肿起;而阔叶箬竹和米箬竹箨鞘的上部包秆较宽松,因而肿起。阔叶箬竹叶片在下表面沿中脉两侧均无纵行的毛茸,而米箬竹一侧密生1纵行的毛茸。形态上的关系也得到本研究结果的支持。
箬竹与箬叶竹相似性系数较高,属于一组;而米箬竹与半耳箬竹属于另一组。无论是箬竹与米箬竹,还是箬叶竹与半耳箬竹,相似性系数都较低,关系也较远。另外,虽然光箨箬竹与粽巴箬竹聚在一组,但是两者的相似性系数仅为0.69,关系也较远。AFLP分析提供的分子证据,并不支持形态学分类(耿伯介等,1996)将米箬竹归入箬竹、半耳箬竹归入箬叶竹、光箨箬竹归入粽巴箬竹中。但目前据此尚不足于否定形态分类的结果,有必要进行进一步的研究。
耿伯介, 王正平. 1996.中国植物志: 第九卷第一分册.北京: 科学出版社.
|
宋红竹, 张绮纹, 周春江. 2007. 杨树部分种的AFLP遗传多样性分析[J]. 林业科学, 43(12): 64-69. |
田清震, 盖钧镒, 喻德跃. 2000. 大豆DNA扩增片段长度多态性(AFLP)研究[J]. 大豆科学, 19(3): 210-217. DOI:10.3969/j.issn.1000-9841.2000.03.003 |
王利, 邢世岩, 王芳, 等. 2008. 银杏雌株种质遗传关系的AFLP分析[J]. 林业科学, 44(4): 48-53. DOI:10.3321/j.issn:1001-7488.2008.04.010 |
王献, 张启祥, 杨秋生, 等. 2005. 利用AFLP研究紫薇的亲缘关系[J]. 北京林业大学学报, 27(1): 59-63. DOI:10.3321/j.issn:1000-1522.2005.01.013 |
王正平. 1997. 中国竹亚科分类系统之我见[J]. 竹子研究汇刊, 16(4): 1-6. |
温太辉. 1991. 关于几个竹亚科分类群的分类问题[J]. 竹子研究汇刊, 10(1): 11-25. |
文亚峰, 谢碧霞, 何钢, 等. 2008. 人心果品种资源亲缘关系的AFLP分析[J]. 林业科学, 44(9): 59-64. DOI:10.3321/j.issn:1001-7488.2008.09.011 |
杨光耀, 赵奇僧. 2000. 苦竹类植物分析及其系统学意义[J]. 西北农业大学学报, 22(4): 551-553. |
朱石麟, 马乃训, 傅懋毅. 1994. 中国竹类植物图志[M]. 北京: 中国林业出版社.
|
Barrett B A, Kidwell K K, Fox P N. 1998. Comparison of AFLP and pedigree-based genetic diversity assessment methods using wheat cultivars from the Pacific Nor thwest[J]. Crop Science, 38(5): 1271-1278. DOI:10.2135/cropsci1998.0011183X003800050026x |
Clayton W D, Renvoize S A. 1986.Genera graminum. Grasses of the world. London: HMSO.
|
Hamilton A J, Lyett G W, Grierson D, et al. 1990. Antisense gene that inhibit synthesis of hormone ethylene in transgenic plants[J]. Nature, 6281(346): 284-287. |
Ohrnberger D. 1999. The bamboos of the world[M]. Elsevier, 44-49.
|
Vos P, Hogers R, Bleeker M, et al. 1995. AFLP-a new technique for DNA fingerprinting[J]. Nucleic Acids Research, 23(21): 4407-4414. DOI:10.1093/nar/23.21.4407 |
Zabeau M, Vos P. 1995. Selective restriction fragment amplication: a general method for DNA fingerprinting. Eumpean Patent Application 92402629.7, publication Number Ep 0534858AI.
|
Zhang Donglin, Dirr M, Price R. 2000. Discrimination and genetic diversity of Cephalotaxus accessions using AFLP markers[J]. J Amer Soc Hort Sci, 125(4 04): 412. |