 2009, Vol. 45
2009, Vol. 45文章信息
- 李海玲, 陈乐蓓, 方升佐, 孙启祥.
- Li Hailing, Chen Lebei, Fang Shengzuo, Sun Qixiang
- 不同杨-农间作模式碳储量及分布的比较
- Comparison of Carbon Storage and Distribution in Different Poplar-Crop Intercropping Patterns
- 林业科学, 2009, 45(11): 9-14.
- Scientia Silvae Sinicae, 2009, 45(11): 9-14.
-
文章历史
- 收稿日期:2008-08-31
-
作者相关文章



2. 山西师范大学 临汾 041004;
3. 中国林业科学研究院 北京 100091
2. Shanxi Normal University Linfen 041004;
3. Chinese Academy of Forestry Beijing 100091
近年来关于农林复合经营系统在固碳及减缓温室效应中作用已越来越多的引起人们的重视(Makundi et al., 2004; Sharrow et al., 2004; Takimoto et al., 2008;Schoeneberger, 2009)。从长远的观点看,农林复合经营比单一经营作物的固碳能力强,据估计,农林复合经营系统中地上部分的碳储量平均在40~150 tC·hm-2之间(IPCC, 2007)。我国学者已开展了陆地碳储量、碳汇及其他与全球变化相关问题的研究,但相对较多的是全国或区域植被或土壤碳方面的研究(王绍强等,2000;方精云等,2001;Fang et al., 2001)。关于农林复合经营系统的研究,目前国内外主要集中在不同农林复合经营模式的水土保持功能、养分循环、光照和水分竞争、立地生产力和经济效益等(裴保华等,2000;王世忠等,2003;方升佐等,2004;许翠等,2006),而有关农林复合经营系统在固碳中的作用研究很少(Oelbermann et al., 2004)。农林复合经营作为1种重要的土地利用类型,在全球关注环境问题、积极寻求增强碳蓄积量、减少碳排放量途径的背景下,研究不同农林复合经营模式的碳储量及其动态具有重要理论和现实意义。本研究选择江苏北部已有大面积栽植的3种典型杨农复合经营模式,以生态系统为整体对不同复合经营模式的碳储量及其动态进行研究,可为准确估算我国碳储量和全球碳循环研究提供基础数据。
1 试验地概况调查地设在宿迁市杨树(Populus deltoids)丰产林生态示范园区,118°18′ E,35°15′ N,濒临洪泽湖、骆马湖,属中纬度暖温带、湿润—半湿润气候区。年平均气温14.9℃,最冷月1月,平均气温为-3 ℃,最热月7月,平均气温为28 ℃,无霜期为210天左右,全年日照时间为2 095.2 h,年降雨量为890~930 mm。土壤为洪泽湖淤积土,表层土有机质含量(1.79±0.27)%,密度(1.41±0.02)g·cm-3,pH为6.60~7.06,全N含量(1.03±0.12)g·kg-1,全P含量0.05%~0.11%,全K含量0.52%~0.78%,地下水位较高,透气透水性较差。
研究选取3种不同的杨农复合经营模式,第1种为宽窄行模式(简称K模式),为4 m×4 m×16 m栽植模式,即每隔16 m栽植4 m×4 m的2行杨树;第2种模式为片林模式(简称P模式),为6 m×10 m栽植模式;第3种模式为小网格模式(简称W模式),中间为面积2 hm2(200 m×100 m)的方形农田,四周南北为3 m×4 m栽植模式,东西为3 m×6 m栽植模式。
试验材料均为‘35杨’(P. deltoids cv. “35”),属美洲黑杨无性系(方升佐等,2004b)。2003年春季植苗造林,苗龄为1年生根1年生干,造林密度K模式为250株·hm-2,P模式为167株·hm-2,W模式为94株·hm-2(其为单位面积上的株数,包括种植农作物的面积在内)。3种模式中间作农作物春季均为小麦(Triticum aestivum),秋季在每中模式中分别设计为水稻(Oryza sativa)、大豆(Glycine max)和玉米(Zea mays)。
2 调查研究方法 2.1 土壤碳的测定和估算2007年3月在3个模式中随机挖取12个土壤剖面(每个模式3个),采集混合土样。按每10 cm一层(其中60~100 cm按20 cm一层)取土壤样品带回实验室,按照国家标准GB 7830-87进行风干、研磨、过筛和混合分样,测定有机碳含量。
100 cm土层碳储量按S=∑(Ci×di×Di×100)计算,式中:i为土壤层次,S为1 m深土层碳储量,t·hm-2;Ci为第i层的土壤的碳含量,g·kg-1;di为第i层的土壤的密度,g·cm-3;Di为第i层的土壤的厚度,cm。
2.2 杨树生物量及各器官的碳含量测定2006年3月,在3种模式林分中分别设置3块固定样地,每年对标准地所有林木的胸径、树高、冠幅进行检测。2007年8月根据林木胸径径阶分布(径阶距为2 cm),在标准地相邻地块选择5个不同径阶的7株标准木伐倒。径阶标准木根据调查样地的林木胸径径阶分布,按权重分配选取,采用全挖法,地上部分按2 m区分段分层切割,分析各个组分(叶、枝、干、皮)生物量分配规律,根系以各伐倒木根株为中心挖取边长为3 m×3 m,深1 m的土壤,按根桩(即骨骼根无分枝部分)、粗根(>0.5 cm)、中根(0.2~0.5 cm)、细根(≤0.2 cm)分级测定地下部分生物量。同时取一定质量的各器官鲜样,在65 ℃恒温箱中烘至恒质量,推算各器官生物量,并分析碳含量。碳含量采用德国Elementar TOC(Germany)元素分析仪测定,每个样品测定3个重复,取平均值。
2.3 农作物生物量及各器官碳含量的测定农作物生物量采用样地单位面积的生物量来估算,设置面积1 m2的正方形样方。根据距树行的距离,按每种农作物在每种间种模式中取6个1 m2的正方形样方,分茎秆、叶、根、穗等全部收获样方中的农作物生物量。取一定量在65 ℃恒温箱中烘至恒质量,推算农作物生物量,并分析碳含量。
2.4 凋落物生物量及碳含量的测定2007年4月在每个标准地中设置5~6个直径为1 m的凋落物收集网,每月收集1次凋落物称量,取一定量在65 ℃恒温箱中烘至恒质量,推算凋落物生物量,并分析碳含量。
2.5 数据处理采用Spss11.0统计分析软件对测定数据进行方差分析(ANOVA)、Duncan检验及回归分析。
3 结果与分析 3.1 3种模式土壤各层碳含量及碳储量土壤碳库在整个生态系统碳库中占有非常重要的地位。土壤中的有机碳主要来源于动植物残体和枯枝落叶经微生物分解转化和化学淋溶的长期过程。3种杨农间作模式土壤各层碳含量及储量的测定结果见表 1。
|
|
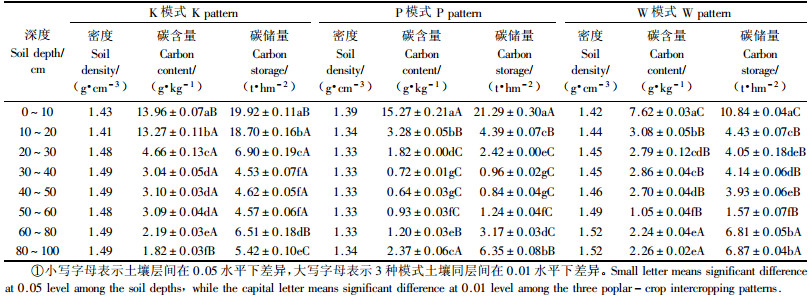
由表 1可以看出:3种模式深度1 m的土壤碳含量变化范围为0.64~15.27 g·kg-1。碳含量在表层土表现相对较高,且有随土壤深度的增加而降低的趋势;其中K模式在20 cm以下碳含量骤减,10~20与20~30 cm 2层之间碳含量差异达2.85倍,而P模式和W模式在土层10 cm处碳含量发生明显变化;3种模式土壤层次间碳含量差异均达到极显著水平。3种模式碳储量均表现出10 cm表层土最大;K模式土壤各层碳沉积能力强于另外两种模式,1 m深度土壤碳储量达71.19 t·hm-2,而P模式和W模式分别为40.67和42.64 t·hm-2,可能是因为K模式杨树栽植密度大,凋落物和根系返还的较多。虽然本研究3种模式所设标准地均处于骆马湖边,土壤同为淤积土,相距不到1 000 m,也可能是土壤异质性存在的所造成。
3.2 杨树不同器官碳含量及其储量 3.2.1 杨树不同器官碳含量分析对3株杨树平均木(胸径为16 cm径阶)取样分析,杨树不同器官碳含量情况如图 1。杨树不同器官碳含量大小顺序为:细根<毛细根<树叶<树枝<粗根<树皮<木材;仅细根和毛细根的碳含量与其他器官存在显著差异,变化范围为459.87~526.70 g·kg-1,变异系数为1.38%~5.47%。

|
图 1 杨树不同器官碳密度的差异 Figure 1 Variation in carbon density at different components of poplar trees |
林木碳储量与林木生物量密切相关,准确测定林木生物量即可估算林木碳储量。根据伐倒的7株不同径阶杨树生物量的实测值,利用林木相对生长法,建立单株林木各器官生物量与林木胸径和树高之间的回归方程如下式:
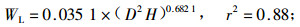
|
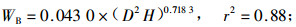
|

|
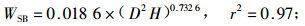
|
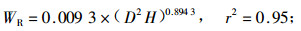
|
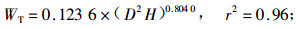
|
式中:WL为叶生物量;WB为枝生物量;WSW为不带皮的树干生物量;WSB为树皮生物量;WR为根生物量;WT为总生物量;D为胸径(cm),H为树高(m),相关显著性水平a=0.01。
杨树林分现存量中各器官的生物量与其相应碳密度之积,即为杨树各器官碳储量,各器官碳储量合计可得林木碳储量。3种模式杨树各器官碳储量见表 2。K,P和W 3种模式各器官生物量大小顺序均是干>枝>根>叶>皮,树干部分生物量占杨树总生物量的比例最大,依次为47.3%,48.4%和48.6%;碳储量在杨树不同器官中的分配与生物量成正比,树干所占比例最高,分别为48.4%,49.5%和49.7%。
|
|
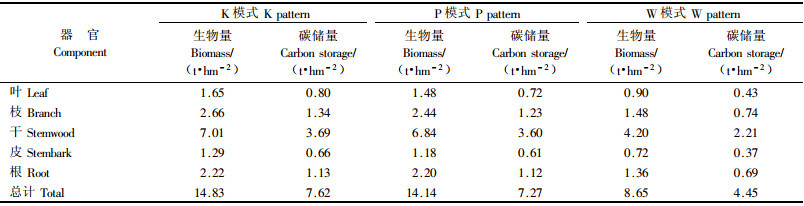
K模式和P模式杨树生物量及碳储量比较接近,而W模式明显低于前2种模式,主要因为W模式杨树配置每公顷只有94株,远低于前2者。
3.3 3种不同模式农作物各器官碳含量及碳储量在3种模式的样地中,对春季作物小麦,秋季作物玉米、大豆、水稻在成熟时分别取样分析,得各作物器官碳含量测定结果及碳储量(表 3)。结果表明:4种作物皆是根的碳含量明显低于其他器官,以水稻根碳含量最小,为(275.70±21.05)g·kg-1,大豆根的碳含量最大,为(424.53±21.69)g·kg-1,小麦和玉米根碳含量接近;4种作物其他器官碳含量差异不显著。
|
|
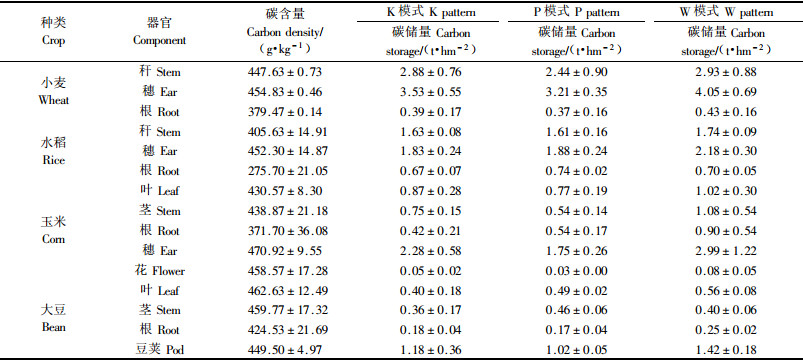
从表 3可以看出:3种模式中春季作物小麦的碳储量都大于秋季3种作物的碳储量,K,P2模式作物收获期碳储量大小为小麦>水稻>玉米>大豆,W模式顺序为小麦>玉米>水稻>大豆。4种作物都是收获部分(穗或豆荚)碳储量所占比例最大,小麦、玉米、大豆均达50%左右,水稻稍低亦达到45%左右。
3.4 3种模式杨树枯落物的碳储量及其动态变化凋落物的碳储量与其生物量密切相关。经测定,调查地枯落物碳含量约为48%(按48%计算)。杨树凋落物的凋落发生在四季,主要有落叶、小枝、苞片和虫粪等其他物质,其中以落叶为主。凋落物凋落年变化出现双峰曲线(图 2),主要集中在9—11月杨树落叶期,但与生长期的初期(4—6月)相比,处于生长旺期的7,8月份凋落物量开始明显增多,这可能由于调查地7,8月降雨较多,地下水位较高,促使叶片凋落;另外由于林冠上层的遮光,下层老叶因为光照不足也会脱落。K,P与W 3种模式含碳量年变化情况都是9月份落叶量最大时,凋落物中含碳量达到最大,全年凋落物含碳量分别为(0.78±0.23),(0.73±0.22)与(0.37±0.09)t·hm-2a-1。

|
图 2 不同间作模式杨树凋落物碳含量的年动态变化 Figure 2 Annual dynamics of carbon content in the litterfall for different poplar-crop intercropping patterns |
杨农复合生态系统碳储量主要包括杨树、春秋季农作物、枯枝落叶和土壤4部分,杨-农系统各模式和各组分碳存量存在差异(图 3)。可以看出:3种模式各组分碳储量均是土壤>农作物>杨树>枯落物,以土壤碳储量占整个生态系统比例最大,达到70%~80%,枯落物碳储量比例最小,只占到0.05%左右。其中,K模式土壤碳储量是生物质碳储量的3.99~4.56倍,P模式土壤碳储量是生物质碳储量的2.41~2.74倍,W模式土壤碳储量是生物质碳储量的2.68~3.42倍。总计各组分碳储量,K,P与W 3种模式不同农作物间作种类(小麦+玉米、小麦+大豆、小麦+水稻)系统碳储量分别为90.02,87.78与89.80 t·hm-2;57.64,56.15与58.24 t·hm-2;60.60,57.15与59.14 t·hm-2。与唐罗忠等(2004)对江苏里下河地区10年生Ⅰ-69杨树人工林的碳储量的估算值(136.2±15.9) t·hm-2相比,本次调查杨农复合生态系统碳储量明显偏低,主要是杨树密度和林龄差异所致。若从单株杨树固定碳的能力来讲,属美洲黑杨无性系的35杨固碳能力并不低,6年生平均达到0.08 tC。

|
图 3 不同间作模式下杨农复合系统的碳储量的差异 Figure 3 Variation in carbon storage of agroforestry system under the three intercropping patterns |
许多学者系统地研究了杨树人工林复合经营体系的结构设计和高效杨树人工林复合经营体系建立的设计原理,总结了现有主要杨树人工林复合经营模式的三维空间配置、潜在生产力、资源利用率、稳定性和经济效益(方升佐等,2004a;Fang et al., 2005;戴晓琴等,2006),本研究主要探讨了3种杨农间作模式的碳储量及分布。研究表明,农林复合经营后,3种模式中6年生杨树的碳储量在4.45~7.62 t·hm-2之间,与单位面积上杨树的株数成正比,但碳储量明显低于10年生的杨树纯林(唐罗忠等,2004;Fang et al., 2007)。
农林复合系统对地上部分植被碳储量的影响,随着时间的推移而变化。与单纯的农田生态系统相比,碳循环周期由原来的1年变为多年,碳截获能力明显增强(主要以木本生物量积累和残留难腐解的枯枝)(Kaur et al., 2000)。本研究区处于我国东部,与我国农田生态系统年碳储量相比,林分6年生时3种模式中作物年碳储量(除K和P模式中的小麦+大豆外)大于农田生态系统作物的最大值9.27 t·hm-2(鲁春霞等,2005)。同时,本研究中土壤表层碳储量与江苏省农田生态系统土壤表层碳储量估算值相比(刘允芬,1998;姜小三等,2005),均持平或高于估算值。这可能是由于研究区紧邻骆马湖以及杨-农复合生态系统杨树的引入栽植,枯枝落叶和根系每年归还到土壤中,从而导致土壤中碳的累积较多,而且有向下层积累的趋势,与李跃林等(2002)的研究结果一致。
从农林复合经营系统的碳储量看,K,P与W 3种模式与不同农作物种类(小麦+玉米、小麦+大豆、小麦+水稻)间作后系统碳储量分别为90.02,87.78与89.80 t·hm-2;57.64,56.15与58.24 t·hm-2;60.60,57.15与59.14 t·hm-2,即系统的碳储量为K模式>W模式>P模式。但作物的碳储量为W模式>K模式>P模式,在林分6年生时,W模式中作物的年碳储量平均比K和P模式分别高出约19.0%和29.5%。同时,4种作物都是收获部分(穗或豆荚)碳储量所占比例最大,小麦、玉米、大豆均达50%左右,水稻稍低亦达到45%左右。由于农作物收获基本上是全量收获地上部分,而其根部所截留的碳所占比例只有5%~17%,这说明每年约有33%~45%的碳流失。因此,应提倡非经济产量资源化利用,如扩大秸秆作为饲料、工业或手工业原料的比例;推行过腹还田、发展沼气(刘允芬,1998);或加强秸秆还田,不但可以有效增加土壤肥力,减少化肥的施用,还可以增加土壤有机碳和作物生物量碳,增加农田的碳截留。
综合现有的研究结果,笔者认为在所研究的3种杨-农间作模式中,以K模式(宽窄行模式,杨树株行距为4 m×4 m×16 m),间作作物为小麦+玉米的农林复合经营系统碳储量最大,可考虑在生产实践中推广应用。但关于在1个轮伐期内杨树生长对农作物产量和生物量的影响,即不同杨农间作模式对农作物碳储量产生影响以及如何延长农林复合经营系统固碳周期(作物残余物的管理)等尚待进一步深入研究。
戴晓琴, 郭兴强, 李鹏, 等. 2006. 平原农区幼龄杨树间作农作物的产量表现[J]. 生态学杂志, 25(12): 1515-1519. DOI:10.3321/j.issn:1000-4890.2006.12.013 |
方精云, 陈安平. 2001. 中国森林植被碳库的动态变化及其意义[J]. 植物学报, 43(9): 967-973. DOI:10.3321/j.issn:1672-9072.2001.09.014 |
方升佐, 徐锡增, 余相, 等. 2004a. 杨-小麦复合经营模式的立地生产力及生态经济效益评价[J]. 林业科学, 40(3): 88-95. |
方升佐, 徐锡增, 吕士行. 2004b. 杨树定向培育[M]. 合肥: 安徽科学技术出版社, 4-28.
|
李跃林, 彭少麟, 赵平, 等. 2002. 鹤山几种不同土地利用方式的土壤碳储量研究[J]. 山地学报, 20(5): 548-552. DOI:10.3969/j.issn.1008-2786.2002.05.007 |
姜小三, 潘剑君, 李学林. 2005. 江苏表层土壤有机碳密度和储量估算和空间分布分析[J]. 土壤通报, 36(4): 501-503. DOI:10.3321/j.issn:0564-3945.2005.04.008 |
刘允芬. 1998. 中国农业系统碳汇功能[J]. 农业环境保护, 17(5): 197-200. |
鲁春霞, 谢高地, 肖玉, 等. 2005. 我国农田生态系统碳蓄积及其变化特征研究[J]. 中国生态农业学报, 13(3): 35-37. |
裴保华, 袁玉欣, 贾玉彬, 等. 2000. 杨农间作光能利用的研究[J]. 林业科学, 36(3): 13-18. DOI:10.3321/j.issn:1001-7488.2000.03.003 |
唐罗忠, 生原喜久雄, 黄宝龙, 等. 2004. 江苏省里下河地区杨树人工林的碳储量及其动态[J]. 南京林业大学学报:自然科学版, 28(2): 1-6. |
王绍强, 周成虎, 李克让. 2000. 中国土壤有机碳及空间分布特征分析[J]. 地理学报, 55(5): 533-543. |
王世忠, 郭浩, 李树民, 等. 2003. 辽西地区几种农林复合型水土保持林模式的研究[J]. 林业科学, 39(3): 163-168. |
许翠, 隋鹏, 谢光辉, 等. 2006. 杨农复合种植模式土壤水分效应及生产力分析[J]. 中国农业科学, 9(4): 758-763. DOI:10.3321/j.issn:0578-1752.2006.04.016 |
Fang J, Chen A, Peng C, et al. 2001. Changes in forest biomass carbon storage in china between 1949 and 1998[J]. Science, 292: 2320-2322. DOI:10.1126/science.1058629 |
Fang S, Xu X, Yu X, et al. 2005. Poplar in wetland agroforestry: A case study of ecological benefit, site productivity, and economics[J]. Wetland Ecology and Management, 13: 93-104. DOI:10.1007/s11273-003-3104-5 |
Fang S, Xue J, Tang L. 2007. Biomass production and carbon sequestration potential in poplar plantations with different management patterns[J]. Journal of Environ mental Management, 85: 672-679. |
IPCC. 2007. 2006IPCC Guidelines for National Greenhouse Gas Inventories.
|
Kaur B, Gupta S R, Singh G. 2000. Soil carbon, microbial activity and nitrogen availability in agroforestry systems on moderately alkaline soils in northern India[J]. Applied Soil Ecology, 15(3): 283-294. DOI:10.1016/S0929-1393(00)00079-2 |
Makundi W R, Sathaye J A. 2004. GHG mitigation potential and cost in tropical forestry—relative role for agroforestry systems[J]. Environ Dev Sust, 6: 235-260. DOI:10.1023/B:ENVI.0000003639.47214.8c |
Oelbermann M, Voroney R P, Gordon A M. 2004. Carbon sequestration in tropical and temperate agroforestry systems: a review with examples from Costa Rica and southern Canada[J]. Agriculture, Ecosystems and Environment, 104: 359-377. DOI:10.1016/j.agee.2004.04.001 |
Schoeneberger M M. 2009. Agroforestry: working trees for sequestering carbon on agricultural lands[J]. Agroforest Syst, 75: 27-37. DOI:10.1007/s10457-008-9123-8 |
Sharrow S H, Ismail S. 2004. Carbon and nitrogen storage in agroforests, tree plantations, and pastures in Westtern Oregon, USA[J]. Agroforest Syst, 60: 123-130. DOI:10.1023/B:AGFO.0000013267.87896.41 |
Takimoto A, Nair P K R, Nair V D. 2008. Carbon stock and sequestration potential of traditional and improved agroforestry systems in the West African Sahel[J]. Agriculture, Ecosystems and Environment, 125: 159-166. DOI:10.1016/j.agee.2007.12.010 |