 2009, Vol. 45
2009, Vol. 45文章信息
- 钟景辉, 张飞萍, 江宝福, 肖梅, 陈家骅.
- Zhong Jinghui, Zhang Feiping, Jiang Baofu, Xiao Mei, Chen Jiahua
- 不同寄主松树对松突圆蚧耐寒性的影响
- Effects of Different Host Pines on the Cold Tolerance of the Pine Scale Insects
- 林业科学, 2009, 45(10): 100-107.
- Scientia Silvae Sinicae, 2009, 45(10): 100-107.
-
文章历史
- 收稿日期:2008-09-18
-
作者相关文章



2. 福建省泉州市森林病虫害防治检疫站 泉州 362000
2. Quanzhou Forest Disease and Pest Control and Quarantine Station of Fujian Province Quanzhou 362000
探讨昆虫的耐寒性对于揭示其地理分布范围、种群波动成因和实施害虫有效控制、益虫充分利用等具有重要意义。昆虫的耐寒性(cold tolerance)与其本身遗传性状有关,也与自身的生理状态和所处的外界环境密切相关。气候的季节性波动、虫体含水量和抗寒物质的积累、冰核物质、地理环境、昆虫世代和发育阶段、低温驯化、食物等均对昆虫耐寒性具有重要影响(Baust,1982;Baust et al., 1985;Zachariassen,1985;Danks,2006;李庆等,2008;景晓红等,2002)。一般而言,昆虫在进入越冬前需积累足够的能量如糖原、脂肪等以满足生存需要(Sakurai et al., 1992),一些昆虫在越冬期间体内的小分子糖类、多元醇类或甘油等的含量相应增加,从而提高冬季的存活机率(Storey et al., 1991),而这些物质的积累与寄主种类和质量密切相关(Zvereva,2002)。如由棉铃虫(Helicoverpa armigera)幼虫取食不同种类植物发育的滞育蛹在水分、糖原、脂质和蛹质量等方面存在显著差异,从而影响其过冷却能力(Liu et al., 2007;杨燕涛等,2003);取食来源于不同地区植物Salix borealis的弧斑叶甲(Chrysomela lapponica)因虫体水分含量不同而致使其越冬存活率显著不同(Zvereva,2002);取食不同种类植物桔小实蝇(Bactrocera dorsalis)蛹的过冷却点和低温暴露存活率也有显著差异,且寄主种类和桔小实蝇发育龄期对该虫耐寒性存在明显的交互作用(任璐等,2006)。这些研究说明:寄主植物的种类和质量不仅影响昆虫的发育、存活、繁殖和世代变化,还对昆虫的越冬、滞育和耐寒性产生显著影响(Tauber et al., 1986;Hunter et al., 1997)。
松突圆蚧(Hemiberlesia pitysophila)是20世纪80年代入侵我国大陆地区的危险性松林害虫,可危害马尾松(Pinus massoniana)、湿地松(P. elliottii)、火炬松(P. taeda)和黑松(P. thunbergii)等16种松树(陈泽藩,1988),现分布于广东、福建和广西等地,引起大面积松林衰退、枯死,造成了严重的生态和经济损失(国家林业局植树造林司等,2005;张星耀等,2003)。从地理纬度看,松突圆蚧的分布目前尚未到达27°N,但吴太平等(1992)通过低温冷冻试验认为该虫分布的地理北界在32—33°N之间,而这一区域内分布着大面积以马尾松、湿地松、火炬松和黑松为主体,多种松属植物构成的松林资源。研究不同寄主松树松突圆蚧的耐寒性,对于该虫的地理分布预测、危害风险分析、种群动态解释和综合治理措施优化等具有重要意义。鉴于此,本文选择马尾松、黑松、湿地松和火炬松上的松突圆蚧为研究对象,通过测定过冷却点和低温暴露试验,分析比较了4种寄主松树上松突圆蚧的耐寒性。
1 材料与方法 1.1 供试虫源寄生于马尾松和湿地松的松突圆蚧虫源均采自福建省泉州市洛江区马甲仙功山景区(25°6.158′N,118°38.255′E附近,海拔368 m),寄生于黑松和火炬松的松突圆蚧虫源均采自福建省南安市罗山国有林场(24°50.979′N,118°19.345′E附近,海拔357 m)。取样时,利用高枝剪剪取虫口密度较多的小枝条,标记后用密封性较好的塑料袋封装,带回实验室后置入人工气候箱沙培保湿隔离培养(25 ℃,RH 70%,16L:8D),持续培养10天以降低因地理环境不同而造成的误差,试验时直接从人工培养箱内选取带有蚧虫的针叶获取虫源。每14天更换新鲜样本1次。
1.2 过冷却点的测定过冷却点测定采用铜-铜镍热电偶进行,通过数据采集器和电脑相连,即时记录测定对象的体温变化数据;低温环境用海尔BCD228-WBSS冰箱(青岛海尔集团)制备,降温速率控制在1 ℃·min-1左右。测定对象为寄生于马尾松、黑松、湿地松和火炬松上的松突圆蚧雌成虫,以及湿地松和马尾松上的2龄性分化后雌、雄若虫。测定时,用挑针从带虫针叶上小心地将松突圆蚧挑下,去除蚧壳,用滤纸将虫体周围的液体吸干,通过透明胶将待测虫体固定在热电偶的探针上,并保证虫体与探针充分接触,并用棉花包裹住带虫探针,放入冰箱内测定其过冷却点值。各寄主、各虫态均重复测定20~71头。过冷却点时间为2008-03—2008-04,此期当地气温逐步回升,但温度波动较大。
1.3 低温暴露试验在测定过冷却点的基础上,选择湿地松和马尾松2种松树上的松突圆蚧作为低温暴露试验的研究对象。由于该虫在福建无明显越冬现象,且世代重叠严重,冬季林间可发现各种虫态(陈顺立等, 2004;童应华等,2006),因此选择初孵若虫、1龄若虫、2龄性分化前若虫、2龄性分化后雌若虫、2龄性分化后雄若虫和雌成虫进行试验。依据自然变温过程和松突圆蚧在12月上中旬(此期福建沿海的最低温约10 ℃左右)种群数量较低的现象(童应华等,2006),设定7种不同强度低温暴露处理,分别为1) 10 ℃处理: 10 ℃×12 h;2)5℃处理: 10 ℃×12 h→5 ℃×6 h;3) 0 ℃处理: 10 ℃×12 h→5 ℃×6 h→0 ℃×6 h;4) -5 ℃处理: 10 ℃×12 h→5 ℃×6 h→0 ℃×6 h→-5 ℃×4 h;5) -10 ℃处理: 10 ℃×12 h→5 ℃×6 h→0 ℃×6 h→-5 ℃×4 h→-10 ℃×4 h;6) -15 ℃处理: 10 ℃×12 h→5 ℃×6 h→0 ℃×6 h→-5 ℃×4 h→-10 ℃×4 h→-15 ℃×4 h;7) -20 ℃处理: 10 ℃×12 h→5 ℃×6 h→0 ℃×6 h→-5 ℃×4 h→-10 ℃×4 h→-15 ℃×4 h→-20 ℃×4 h;另设对照(25 ℃,RH 70%,16L:8D处理)。其中5 ℃以上温度环境(包括5 ℃)由PYX-1250Q-A型号人工气候箱(广东韵关科力试验仪器有限公司)制备;0 ℃以下温度环境(包括0 ℃)由海尔BCD228-WBSS冰箱制备。
低温暴露处理时,分别从2种寄主植物上随机抽取8×100束带虫针叶(每100束针叶可保证松突圆蚧各虫态个体数量大于30头),将每100束针叶用1层废报纸包扎作为1个处理对象,2种寄主植物共计8组(16个)处理对象。将各处理对象分别进行7种低温暴露和1种对照处理,结束后将各处理对象取出并直接放入设置好的人工气候箱内(25 ℃,RH 70%,16L:8D)。待最后1种处理(-20 ℃处理)结束并在相同条件的人工气候箱内恢复24 h后,将对照和各处理的针叶取出,解剖镜下检查并统计松突圆蚧各虫态的死亡和存活个体数。存活标准:去除蚧壳后虫体饱满、色泽光亮;低温致死标准:虫体皱缩、体色棕褐色或带有深色斑点、斑带;虫体干瘪、残缺,蚧壳内无虫、蚧壳上有寄生蜂羽化孔视为其他原因致死,不作统计。试验设3重复。试验时间与1.2相同。
1.4 数据分析 1.4.1 过冷却点分析采用方差分析(ANOVA)结合新复极差法(Duncan法)比较各地区松突圆蚧过冷却点。过冷却点频次分布的正态性采用χ2法检验。
1.4.2 耐寒性评价Nedvěd等(1998)和Ma等(2006)应用双变量逻辑斯谛方程评价昆虫的耐寒性。方程如下:
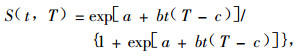
|
(1) |
式中:S为死亡率,t为低温暴露时间,T为低温暴露温度,a,b和c为常数;其中c的生物学意义为对昆虫造成冷伤害的上限温度(the upper limit of chill injury zone, ULCIZ),-a/b的生物学意义为造成50%昆虫个体死亡的有效伤害低温累积(semi-lethal sum of injurious temperature, LSIT50)(景晓红等,2004)。
方程(1)充分考虑了低温暴露强度和低温暴露时间2个因素对昆虫死亡的交互影响;然而,该方程只适用于恒定低温暴露试验中昆虫耐寒性的评价;事实上,自然条件下昆虫经历的低温环境是一个逐渐降温的过程,其死亡情况是对变化低温伤害累积的反应,因此在低温暴露试验中设置逐渐降温的低温环境,无疑更为贴近自然实际情况,从而提高评价昆虫耐寒性的准确度。
为了评价变化低温暴露试验中昆虫的耐寒性,对方程(1)进行如下改进:

|
(2) |
式中:S为死亡率,ti为特定低温暴露时间,其单位可根据试验具体情况选择日、小时或分钟等时间单元,Ti为特定低温暴露温度,单位为℃,i为变化温度处理数,i=1,2,3,…,∑tiTi为变化低温处理的低温累积;a,b和c为常数,其中c的生物学意义为对昆虫造成冷伤害的有效低温累积上限(the upper limited sum of chill injurious temperature, ULSCIT),-a/b的生物学意义为造成50%昆虫个体死亡的有效伤害低温累积(LSIT50)。式中∑tiTi可以通过各温度下暴露的时间计算,S可通过试验测定,a,b,c 3个参数可通过麦夸特法或加速单纯形法等方法估算。
本文采用改进的方程(2)评价不同寄主松树松突圆蚧的耐寒性,采用加速单纯形法估算各参数。然而,由于一些昆虫可能在0 ℃或0 ℃以上低温时即造成伤害,而在0 ℃条件下的低温累积无法反应温度和时间的交互作用,因此在设置的温度大于或等于0 ℃时,需对所设置的各处理温度进行重新赋值才能反映试验的时间×温度处理情况。本文将10℃条件赋值为-1,在此基础上每下降1 ℃,其赋值相应减1,具体为:令T(10 ℃)=-1,T (5 ℃)= -6,T(0 ℃)=-11,T(-5 ℃)=-16,T (-10 ℃) =-21,T(-15 ℃)=-26,T (-20 ℃)= -31。采用这一方法拟合方程(2),据此计算LSIT50,得出的结果只用于比较不同寄主松树松突圆蚧的耐寒性。死亡率采用各重复中校正死亡率的平均值代替:校正死亡率=(处理组死亡率-对照组死亡率)/(1-对照组死亡率)。
2 结果与分析 2.1 不同寄主松树松突圆蚧的过冷却点松突圆蚧过冷却点测定结果见表 1。不同寄主间雌成虫的过冷却点具有显著差异(F=11.700, df=3, 200, P=0.000 0),寄生黑松和马尾松雌成虫的过冷却点比寄生湿地松和火炬松的显著较高(P<0.05),2类过冷却点平均值相差0.9~2.3 ℃,但寄生黑松和马尾松的雌成虫之间、寄生湿地松和火炬松的雌成虫之间未表现显著差异(P>0.05);寄生马尾松和湿地松2龄性分化后雌若虫的过冷却点也具有显著差异(F=13.838, df=1, 62, P=0.000 4),前者的平均值比后者高2.08 ℃(P<0.05),但2种寄主间2龄性分化后雄若虫的过冷却点无显著差异(F=0.943, df=1, 45, P=0.336 6)。总体上,各寄主松树松突圆蚧过冷却点的平均值波动在(-15.20±0.42)~(-12.91±0.13) ℃之间,个体值则波动在-21.8~-8.8 ℃之间。从过冷却点测定的极值看(表 1),寄生马尾松的雌成虫、寄生湿地松的雌成虫和2龄性分化后雌若虫过冷却点值差别较明显,个体间差异较大。
|
|

χ2拟合优度检验表明:各寄主、各虫态的过冷却点均服从正态分布(P>0.05)。以1 ℃为间距,绘制不同寄主松树松突圆蚧3种虫态过冷却点值的频次分布图(图 1,2和3)。对于雌成虫,寄生湿地松的过冷却点频次分布最分散,大多数个体的过冷却点分布在-20~-12 ℃之间,2个不太明显的峰值分别出现在-18~-16 ℃之间和-13~-12 ℃之间(图 1B);寄生马尾松的过冷却点频次分布也较分散,但与湿地松相比,其分布相对更集中,多数个体的过冷却点集中在-15~-11 ℃之间,并呈明显的单峰分布,其峰值出现在-15~-13 ℃之间(图 1C);寄生黑松(图 1A)和火炬松(图 1D)的过冷却点频次分布范围则均明显较窄,前者的多数个体集中分布于-14~-12 ℃之间,后者集中分布于-15~-13 ℃之间(图 1D),二者也均呈明显的单峰分布。

|
图 1 不同寄主松树松突圆蚧雌成虫过冷却点的频次分布 Figure 1 Frequency distribution of SCP for H. pitysophila adult females on different pines A,B,C,D:黑松、湿地松、马尾松和火炬松 A, B, C, D meansP. thunbergii, P. elliottii, P. massoniana and P. taeda, respectively. |

|
图 2 不同寄主松树松突圆蚧2龄性分化后雌若虫过冷却点的频次分布 Figure 2 Frequency distribution of SCP for H. pitysophila 2nd instar female nymph after sexual differentiation on different pines A,B:马尾松和湿地松P. massoniana and P. elliottii, respectively.下同The same below. |

|
图 3 不同寄主松树松突圆蚧2龄性分化后雄若虫过冷却点的频次分布 Figure 3 Frequency distribution of SCP for H. pitysophila 2nd instar male nymph after sexual differentiation on different pines |
对于2龄性分化后雌若虫(图 2),寄生马尾松的过冷却点频次分布幅度明显大于湿地松,前者出现3个峰值,分别为-20~-18 ℃,-16~-15 ℃和-14~-13 ℃之间(图 2A),后者则集中分布于-16~-13 ℃之间,并呈单峰分布,峰值出现在-14~-13 ℃之间(图 2B)。对于2龄性分化后雄若虫,寄生马尾松的过冷却点频次分布幅度则明显小于湿地松,前者波动在-16~-8 ℃之间(图 2A),多数个体集中分布于-16~-13 ℃之间(图 3A),后者波动在-21~-10 ℃之间,多数个体集中于-16~-10℃之间(图 3B)。
2.3 低温暴露下不同寄主松树松突圆蚧的死亡率不同寄主松树松突圆蚧各虫态低温暴露下死亡率情况见图 4。从图 4可看出,在10~-20 ℃之间,寄生马尾松和湿地松的松突圆蚧各虫态的死亡率均随着暴露温度降低而升高。然而,不同寄主松树间表现出明显差异。从曲线变动趋势看,寄生马尾松和湿地松的相同虫态对低温暴露致死反应的敏感温度范围(死亡率急剧增大的温度范围)不同,前者的初孵若虫、1龄若虫、2龄性分化前若虫、2龄性分化后雌若虫、2龄性分化后雄若虫、雌成虫和种群总体的低温暴露致死反应敏感温度范围分别在-20~-5 ℃,-20~-10 ℃,-15~-5 ℃,-15~-5 ℃,-15~-10 ℃,-15~-5 ℃和-15~-5 ℃之间,而后者分别在-20~10 ℃,-20~5 ℃,-20~5 ℃,-15~0 ℃,-5~0 ℃,-20~0 ℃和-20~5 ℃之间,可见寄生马尾松的松突圆蚧各虫态和种群总体对低温暴露致死反应的起始敏感致死低温基本均比寄生湿地松的低。此外,在-20 ℃低温暴露下,寄生马尾松的松突圆蚧各虫态和种群总体的死亡率也均比寄生湿地松的低。

|
图 4 不同寄主松树松突圆蚧低温暴露下死亡率曲线 Figure 4 Mortality curve of H. pitysophila on different pines under exposure to low temperature Ⅰ~Ⅶ分别指初孵若虫、1龄若虫、2龄性分化前若虫、2龄性分化后雌若虫、2龄性分化后雄若虫、雌成虫和种群总体。表 2与此相同。 Ⅰ~Ⅶ are refered to the pest developing stage of newly-hatched nymph, 1st instar nymph, 2nd instar nymph before sexual differentiation, 2nd instar female nymph after sexual differentiation, 2nd instar male nymph after sexual differentiation, adult females and population, respectively. Same for the following Tab. 2. |
|
|

不同寄主松突圆蚧各虫态低温暴露死亡率双变量逻辑斯谛方程和致死中有效伤害低温累积见表 2。从表 2可以看出,各方程的决定系数(R2)基本均大于0.9,P值均小于0.05,说明各方程拟合效果好,寄生2种植物的松突圆蚧各虫态和种群总体的低温暴露死亡率变化规律均符合改进后的双变量逻辑斯谛曲线模型(方程2)。比较不同寄主松突圆蚧相同虫态的致死中有效伤害低温累积(LSIT50)可知,除雌成虫外,寄生马尾松的松突圆蚧各虫态及种群总体的LSIT50均明显比寄生湿地松的相应虫态和种群总体更低。从表还可看出,寄生马尾松的松突圆蚧以初孵若虫的LSIT50最低,而寄生湿地松的则以雌成虫的LSIT50最低,这说明寄主松树与松突圆蚧发育龄期对其耐寒性可能存在交互作用。
3 小结与讨论过冷却点是衡量昆虫耐寒性的一个必要指标,并被认为是昆虫生存环境的低温极限值。但一些昆虫在高于平均过冷却点的温度下大量死亡,而另一些昆虫则在低于平均过冷却点的温度下仍有较高的存活率(Baust et al., 1985)。这与不同种类昆虫的抗寒策略和过冷却点个体间的巨大差异有关,也与昆虫过冷却点易受其他因子影响而出现的不确定性有关。一些研究证实:昆虫的生理状态和所处环境的气温变化过程与其过冷却点密切相关,而寄主植物对植食性昆虫的含水量、体质量、能量储备、发育速率以及小分子抗寒性物质等均具有重要影响,因而被认为是影响过冷却点的一个重要因子(Sakurai et al., 1992;Zvereva,2002;Liu et al., 2007)。不同寄主松树间松突圆蚧雌成虫、2龄性分化后雌若虫的过冷却点出现不同程度的显著差异(表 1),而2龄性分化后雄若虫之间未现显著差异,这一结果既为上述观点提供了支持,也说明寄主植物对植食性昆虫过冷却点的影响与昆虫本身的发育阶段和性别等有关。然而,由于昆虫抗寒策略的差异和过冷却点的不确定性,一些观点认为过冷却点并非昆虫耐寒性评价足以信赖的指标(景晓红等,2004)。
昆虫在低温暴露下的死亡率被认为是一种比过冷却点更可信赖的耐寒性评价指标,通常采用单变量Logistic非线性回归、双变量Logistic非线性回归或直线回归等模型进行综合评价(景晓红等,2004;Nedvěd et al., 1998;Ma et al., 2006;Régnièrea et al., 2007),然而这些评价方法只能对恒定低温暴露下的昆虫耐寒性进行评价。自然条件下,昆虫受到的低温伤害往往是逐渐下降的低温环境与暴露时间交互作用的结果,因此通过逐渐降温的低温暴露试验测定昆虫的耐寒性无疑可以获得更接近自然的结果,但目前尚无合适的评价方法或模型。本文对Nedvěd等(1998)提出的双变量Logistic非线性方程进行改进(见1.4.2部分),并利用改进后的模型分析7种不同强度的变化低温暴露试验中寄生马尾松和湿地松的松突圆蚧各虫态的死亡率与低温的关系。结果显示,2种寄主松树上的松突圆蚧各个虫态以及种群总体均符合改进后的双变量Logistic模型,可见该模型适用于变化低温暴露试验中松突圆蚧耐寒性评价。通过该模型计算2种寄主各虫态的致死中有效伤害低温累积(LSIT50)表明:除雌成虫外,寄生马尾松的松突圆蚧各虫态及种群总体的LSIT50均明显比寄生湿地松的相应虫态和种群总体更低,这说明寄主松树对松突圆蚧的耐寒性造成了影响,该虫湿地松种群的耐寒性比马尾松种群弱。此外,2种寄主松树松突圆蚧雌成虫过冷却点的比较结果与低温暴露的LSIT50一致,而2龄性分化后雌、雄若虫则不一致,这说明过冷却点仅能作为评价该虫耐寒性的一个参考指标,也说明昆虫种群的耐寒机制是一个复杂的问题。
寄主植物对昆虫耐寒性的影响是一个十分复杂的过程。寄主植物一方面通过影响昆虫体内的糖元、脂质、水分、甘油和小分子抗寒物质等的含量以及与耐寒有关的生理代谢过程而影响其耐寒性,另一方面也可以通过影响昆虫的发育进度,调节昆虫进入冬季的虫态而影响其越冬存活率(Tauber et al., 1986;Hunter et al., 1997;Zvereva,2002;Liu et al., 2007;杨燕涛等,2003)。从本文看,不同种类松树对松突圆蚧的过冷却点和低温暴露生存情况均产生了明显影响,然而有关这一影响形成的机制仍需进一步研究。松树种类与松突圆蚧发育龄期对耐寒性可能存在交互作用,这与任璐等(2006)对桔小实蝇的研究结果相近,这一现象是否具有普遍性仍需进一步验证。对福建和广东的观察表明:松突圆蚧在湿地松上的种群数量明显低于马尾松(张星耀等,2003),这与2种植物上松突圆蚧的耐寒性相一致,在一定程度上体现了该虫的适生特性,而松突圆蚧耐寒性与寄主松树抗虫性之间的关系也是一个值得深入研究的问题。
此外,松突圆蚧的巨大危害性和几乎囊括中国长江以南所有松属植物的寄主范围使其成为该区域最为危险的松林检疫害虫之一。目前,该虫仅分布于广东、福建、广西和日本冲绳等地,从地理纬度上看尚未到达27°N,而我国37°N以南分布着大面积该虫的适生寄主,且这些区域平常年份的极端低温基本不超过-20 ℃,但是无论寄生马尾松还是湿地松,松突圆蚧在-20 ℃条件下均保持一定的种群存活率(图 4)。因此初步认为,我国长江以南区域均为松突圆蚧的潜在入侵区域。
陈顺立, 武夫华, 侯沁文. 2004. 松突圆蚧生物学特性研究[J]. 福建林业科技, 31(2): 1-4. DOI:10.3969/j.issn.1002-7351.2004.02.001 |
陈泽藩. 1988. 十五种松树对松突圆蚧抗性的初步研究[J]. 森林病虫通讯, (2): 1-2. |
国家林业局植树造林司, 国家林业局森林病虫害防治总站. 2005. 中国林业检疫性有害生物及检疫技术操作办法[M]. 北京: 中国林业出版社, 1-6.
|
景晓红, 康乐. 2004. 昆虫耐寒性的测定与评价方法[J]. 昆虫知识, 41(1): 7-10. DOI:10.3969/j.issn.0452-8255.2004.01.002 |
景晓红, 康乐. 2002. 昆虫耐寒性研究[J]. 生态学报, 22(12): 2202-2207. DOI:10.3321/j.issn:1000-0933.2002.12.026 |
李庆, 王思忠, 封传红, 等. 2008. 西藏飞蝗(Locusta migratoria tibetensis Chen)耐寒性理化指标[J]. 生态学报, 28(3): 1314-1320. DOI:10.3321/j.issn:1000-0933.2008.03.048 |
任璐, 陆永跃, 曾玲, 等. 2006. 寄主对桔小实蝇耐寒性的影响[J]. 昆虫学报, 49(3): 447-453. DOI:10.3321/j.issn:0454-6296.2006.03.016 |
童应华, 陈顺立, 张飞萍. 2006. 松突圆蚧种群动态及与气象因子的关系[J]. 福建林学院学报, 26(2): 107-110. DOI:10.3969/j.issn.1001-389X.2006.02.003 |
吴太平, 周昌清. 1992. 松突圆蚧冷冻试验及其潜在地理分布范围北界的推测[J]. 生态科学, (1): 62-66. |
杨燕涛, 谢宝瑜, 高增祥, 等. 2003. 寄主植物对棉铃虫越冬蛹抗寒能力的影响[J]. 昆虫知识, 40(6): 509-512. DOI:10.3969/j.issn.0452-8255.2003.06.007 |
张星耀, 骆有庆. 2003. 中国森林重大生物灾害[M]. 北京: 中国林业出版社, 256-275.
|
Baust J G, Rojas R R. 1985. Review-insect cold hardiness: Facts and fancy[J]. Journal of Insect Physiology, 31: 755-759. DOI:10.1016/0022-1910(85)90067-8 |
Baust J G. 1982. Environmental triggers to cold hardening[J]. Comparative Biochemistry and Physiology A, 73: 563-570. DOI:10.1016/0300-9629(82)90263-8 |
Danks H V. 2006. Insect adaptations to cold and changing environments[J]. Canadian Entomology, 138: 1-23. DOI:10.4039/n05-802 |
Hunter M D, McNeil J N. 1997. Host-plant quality influences diapause and voltinism in a polyphagous insect herbivore[J]. Ecology, 78: 977-986. DOI:10.1890/0012-9658(1997)078[0977:HPQIDA]2.0.CO;2 |
Liu Z D, Gong P Y, Wu K J, et al. 2007. Effects of larval host plants on over-wintering preparedness and survival of the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae)[J]. Journal of Insect Physiology, 53: 1016-1026. DOI:10.1016/j.jinsphys.2007.05.005 |
Ma R Y, Hao S G, Tian J, et al. 2006. Seasonal variation in cold-hardiness of the japanese pine sawyer Monochamus alternatus (Coleoptera: Cerambycidae)[J]. Environmental Entomology, 35(4): 881-886. DOI:10.1603/0046-225X-35.4.881 |
Nedvěd O, Lavy D, Verhoef H A. 1998. Modeling time-temperature relationship in cold injury and effect of high temperature interruptions on survival in a chill sensitive Collembolan[J]. Functional Ecology, 12: 816-824. DOI:10.1046/j.1365-2435.1998.00250.x |
Régnièrea J, Bentz B. 2007. Modeling cold tolerance in the mountain pine beetle, Dendroctonus ponderosae[J]. Journal of Insect Physiology, 53: 559-572. DOI:10.1016/j.jinsphys.2007.02.007 |
Sakurai H, Kawai T, Takeda S. 1992. Physiological changes related to diapause of the lady beetle, Harmonia axyridis (Coleoptera: Coccinellidae)[J]. Applied Entomology and Zoology, 27: 479-487. DOI:10.1303/aez.27.479 |
Storey K B, Storey J M. 1991. Biochemistry of cryopretectants//Lee R E, Delinger D L. Insect at Low Temperature. New York: Chapman & Hall, 64-93.
|
Tauber M J, Tauber C A, Masaki S. 1986. Seasonal Adaptations of Insects[M]. New York: Oxford University Press.
|
Zachariassen K E. 1985. Physiology of cold tolerance in insect[J]. Physiological Review, 65: 977-832. |
Zvereva E L. 2002. Effects of host plant quality on overwintering success of the leaf beetle Chrysomela lapponica (Coleoptera: Chrysomelidae)[J]. European Journal of Entomology, 99: 189-195. DOI:10.14411/eje.2002.027 |