 2009, Vol. 45
2009, Vol. 45文章信息
- 麦苗苗, 石大兴, 王米力, 李伟.
- Mai Miaomiao, Shi Daxing, Wang Mili, Li Wei
- PEG处理对连香树种子萌发与芽苗生长的影响
- Seed Germination and Seedling Growth of Cercidiphyllum japonicum with PEG Treatment
- 林业科学, 2009, 45(10): 94-99.
- Scientia Silvae Sinicae, 2009, 45(10): 94-99.
-
文章历史
- 收稿日期:2008-11-19
-
作者相关文章



连香树(Cercidiphyllum japonicum)为连香树科连香树属落叶乔木,是东亚特有的珍稀濒危树种,属国家二级保护稀有种(傅立国,1991)。连香树起源古老,是第三纪古热带植物的孑遗种植物,作为具有造纸、香料、药用和木材等多种经济效益及绿化、美化、优化环境等生态效益的植物资源,具有较高的科研、开发和利用价值。近年来,连香树作为彩色叶树种已被世界各国广泛引种栽培保护(Christophe et al., 1996; Gibbons,1995)。自从该种被发现以来,人们对连香树在生物学和生态学特性、形态解剖、系统分类、木材结构、植物化学、引种繁殖及遗传多样性等方面进行了研究,但有关连香树抗旱性的研究还尚未见报道。种子是植物最重要的繁殖材料,它在发芽阶段的耐旱状况在一定程度上反映了该树种的耐旱程度,种子发芽状况也是判定种子质量、确定播种量的一项重要指标。本文应用高分子渗透调节剂聚乙二醇(polyethylene glycol,简称PEG)处理连香树种子,对其发芽能力、生理变化及抗旱能力进行研究,以期为抗旱机制和抗旱品种选育提供参考和依据。
1 材料与方法 1.1 材料试验用连香树种子2006年10月采自四川宝兴赶羊沟境内约40年株龄的2棵母株,采后种子保存于-20 ℃冰箱中。试验于2008年7月在四川农业大学林木育种实验室进行。
1.2 方法采用不同质量浓度的聚乙二醇(PEG 6000)溶液模拟渗透胁迫,其质量浓度分别为95,140,170,221,250,301和345 g·L-1,与之相对应的溶液水势约为-0.2,-0.3,-0.5,-0.7,-0.9,-1.1和-1.3 MPa(曹帮华,2005)。
1.2.1 PEG引发处理先将连香树种子用福尔马林消毒10 min,再用蒸馏水清洗数次,然后在25 ℃室温下,把种子分别浸泡在不同质量浓度的PEG溶液中,以蒸馏水作对照,浸种48 h。每隔3 h摇动试管几次,以利于通气。再将种子均匀摆入铺有2层无菌滤纸的发芽床中,而后用蒸馏水渗透发芽床,每床摆放100粒,重复3次。温度25 ℃,光照2 000 lx,16 h光照培养。
1.2.2 PEG胁迫处理先将连香树种子用福尔马林消毒10 min,再用蒸馏水清洗数次后直接播种。将种子均匀摆入铺有2层无菌滤纸的发芽床中,而后用不同质量浓度的PEG溶液渗透发芽床,以蒸馏水作对照,每床摆放100粒,重复3次,每隔24 h为各处理发芽床用称量法补加水分至原质量,以保持恒定水势。温度25 ℃,光照2 000 lx,16 h光照培养。
1.2.3 萌发及芽苗形态指标种子萌发第20天统计发芽率,30天统计存活率。发芽率=种子发芽数/供试种子数×100%。存活率=存活芽苗数/总芽苗数×100%。发芽试验结束后,在每个处理的3次重复中分别随机抽取芽苗20株,测定其芽苗的胚轴和胚根长度。
1.2.4 生理生化指标可溶性蛋白含量的测定采用Bradford法(Bradford,1976),可溶性糖含量的测定采用蒽酮法(张志良,2003),脯氨酸含量的测定采用磺基水杨酸法(张殿忠等,1990),丙二醛(MDA)含量参考赵世杰等(1994)采用的测定法,超氧化物歧化酶(SOD)活性的测定采用NBT光照化学还原法(Dhindsa et al., 1981),过氧化物酶(POD)活性的测定采用愈伤木酚氧化法(李合生等,2001),过氧化氢酶(CAT)活性参考叶功富等(1999)采用的测定法。
1.3 数据分析采用SAS 9.1.3软件对种子萌发、芽苗生长及生理生化等指标进行单因素方差分析(one-way ANOVA),采用LSD法检验其差异显著性,结果以平均值±标准差(Mean±SD)表示。
2 结果与分析 2.1 PEG预处理和干旱胁迫对种子萌发过程的影响PEG预处理对种子萌发的影响。同对照相比,不同质量浓度PEG溶液预先浸种对连香树种子萌发和芽苗生长的变化趋势相似。伴随PEG质量浓度的增加,萌发率和苗长(包括胚轴和胚根)均先升高后降低(表 1)。PEG质量浓度为140~301 g·L-1时都能有效提高连香树种子的萌发率,其中以221 g·L-1为最好,萌发率为68.67%;同时,芽苗的胚轴和胚根长度也明显增加,在221 g·L-1时分别为1.70和1.80 cm。综合种子萌发和芽苗生长分析,221 g·L-1质量浓度预处理连香树种子效果较明显,不仅萌发率、胚轴长和胚根长均达到最大值,而且萌发率与其他处理差异极显著,胚根与其他处理差异显著。在种子萌发期间,没有出现芽苗死亡的现象,各处理存活率等同于萌发率。
|
|
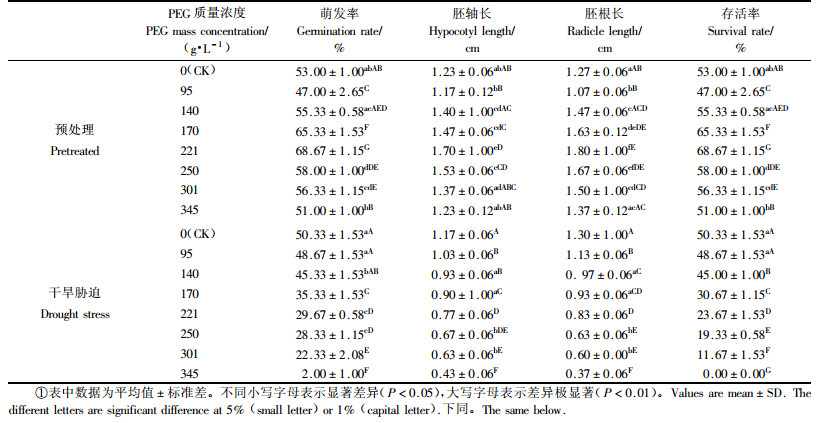
PEG干旱胁迫对种子萌发的影响。从表 1可见,随PEG质量浓度的升高,连香树种子的萌发率、存活率、胚轴和胚根长度呈明显的下降趋势,胚轴和胚根长度都逐渐变短。在不添加PEG的处理下,各项萌发和生长指标均为最高值,种子萌发率和芽苗存活率均为50.33%,除与95 g·L-1差异不显著外,与其他处理差异显著。此时芽苗长势较好,胚轴和胚根长度分别为1.17和1.30 cm,均与其他处理差异极显著。随着水势的下降和胁迫的加重,种子萌发受到抑制,且芽苗胚根根尖变黑,有坏死现象出现。胁迫越严重,芽苗胚根坏死部分越多,死亡数量越多,存活率越低,尤其从221 g·L-1质量浓度开始变为明显。在较高PEG质量浓度(345 g·L-1)下,连香树种子的萌发率仅为2.00 %,胚轴和胚根长度分别仅有0.43和0.37 cm,没有芽苗存活下来,表明在较高渗透势下,芽苗受到的干旱胁迫较为严重,生理代谢紊乱并难以恢复,最终导致死亡。
2.2 PEG预处理和干旱胁迫对芽苗生理生化指标的影响在PEG干旱胁迫试验中,质量浓度为250~345 g·L-1时种子萌发率和芽苗存活率较低、芽苗长势较差,没有进行相关生理生化指标测定,其他处理测定结果详见表 2,3。
|
|

|
|
可溶性蛋白具有较强的亲水胶体性质,可影响细胞的保水力。经PEG预处理后,随着PEG质量浓度的增大,可溶性蛋白含量呈现出增加的趋势。不同质量浓度的PEG处理均明显高于对照,除质量浓度95与140 g·L-1差异显著、质量浓度221与250 g·L-1差异不显著外,其他质量浓度间差异极显著。对于刚刚萌发的种子而言,可溶性蛋白的含量从某种程度上反映了其代谢活性的高低,这一结果从侧面说明PEG预处理能促进萌发中种子活性的提高。由表 2可知,在PEG处理模拟干旱胁迫下,随着PEG质量浓度的增加,连香树芽苗可溶性蛋白含量先增加后下降。当PEG质量浓度在95,140 g·L-1时,含量分别为25.63,28.20 mg·g-1,均高于对照; 当质量浓度为221 g·L-1时含量较低,仅为19.70 mg·g-1,低于对照值24.67 mg·g-1。除质量浓度95 g·L-1与对照差异不显著外,不同质量浓度PEG的连香树芽苗中可溶性蛋白含量的差异极显著。
2.2.2 对可溶性糖含量的影响可溶性糖是呼吸底物,在渗透调节过程中,呼吸底物消耗,可溶性糖含量下降; 呼吸作用加强,可溶性糖含量上升。从表 2可见,2组处理的可溶性糖含量都随着PEG质量浓度的增大而呈现出增加的趋势,均高于对照,以维持体内的渗透压。可溶性糖含量随PEG质量浓度的增加而增加,表明PEG能促进种子内部的贮藏多糖向可溶性糖含量的转化并加速糖异生过程(李合生等,2002)。PEG质量浓度在345 g·L-1时,预处理后种子的可溶性糖含量最高(8.80 mg·g-1),为对照值的2.08倍,不同质量浓度的PEG浸种处理均与对照差异极显著; 干旱胁迫下质量浓度为221 g·L-1时,种子的可溶性糖含量最高(9.37 mg·g-1),为对照值的2.16倍,除质量浓度95与140 g·L-1差异不显著外,不同质量浓度PEG的连香树芽苗中可溶性糖含量的差异显著。
2.2.3 对脯氨酸含量的影响脯氨酸含量的提高是逆境条件下植物的自卫反应之一,细胞脯氨酸含量增加,维持了细胞的膨压,同时可以保护酶和膜系统免受毒害(刘奕清,2008)。在PEG预处理下随着PEG质量浓度的增加,连香树芽苗中脯氨酸含量逐渐升高,均高于对照,质量浓度为345 g·L-1时达到最高值45.13 μg·g-1,为对照值的1.61倍。由表 2可知,干旱胁迫下,PEG质量浓度在95,140 g·L-1时,连香树芽苗脯氨酸含量较低,分别为17.63,21.53 μg·g-1,含量均低于对照值22.67 μg·g-1; 质量浓度在170,221 g·L-1时,脯氨酸含量分别为33.60,38.37 μg·g-1,均明显高于对照。除140 g·L-1与对照差异不显著外,不同质量浓度PEG的连香树芽苗中脯氨酸含量的差异极显著。当胁迫达到一定程度时会导致植物体内游离脯氨酸的积累,进行渗透调节,以适应水分胁迫环境。在本试验中,质量浓度在170 g·L-1时含量急剧上升,表明芽苗出现了适应干旱胁迫的渗透调节反应,质量浓度在95~140 g·L-1时有利于种子萌发后的生长,质量浓度170~221 g·L-1则抑制生长,已经对种子产生了胁迫作用。
2.2.4 对丙二醛(MDA)含量的影响MDA是膜脂过氧化的最终产物,已被证明MDA的积累来自不饱和脂肪酸的降解,它是由体内自由基引发而产生的。因此,MDA的积累在一定程度上反映了体内自由基的活动状态,MDA积累多,说明超氧阴离子与羟基自由基处于较高水平。从表 2可见,经PEG预处理种子后,MDA含量随着PEG质量浓度的增大而呈现出降低的趋势,均低于对照,MDA含量峰值比对照减少了1.51%。8个PEG质量浓度预处理后MDA含量缓慢降低,这表明PEG处理能降低膜脂过氧化反应,有利于膜系统损伤的修复。各处理均与对照差异极显著。PEG干旱胁迫下种子的MDA含量随着PEG质量浓度的增大而呈现出增加的趋势,MDA含量在质量浓度为95~170 g·L-1时缓慢升高,说明连香树对水分胁迫较为敏感,轻微质量浓度就已经开始出现不适,但总的来说MDA含量的增幅不大,表明其受到伤害程度较小; 质量浓度在221 g·L-1时MDA含量急剧升高,表明连香树受到严重伤害。在干旱胁迫下各处理差异极显著。
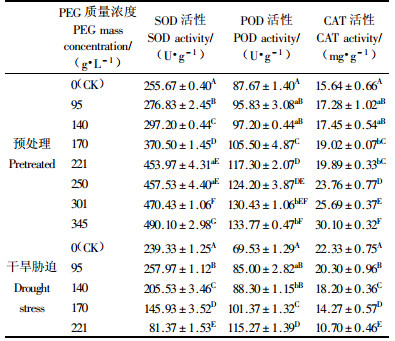
2.2.5 对超氧化物歧化酶(SOD)活性的影响SOD的主要功能是清除氧自由基,是防止其对细胞膜系统伤害的一种很重要的抗氧化酶。从表 3可以看出,随着PEG预处理质量浓度的增加,连香树芽苗中SOD的活性逐渐升高,均高于对照,在345 g·L-1质量浓度时达到最高值490.10 U·g-1,为对照值的1.92倍。除质量浓度221与250 g·L-1差异不显著外,其他处理差异极显著。在不同PEG质量浓度干旱胁迫处理中,SOD活性都呈现先升后降的变化趋势。连香树芽苗中的SOD活性在95 g·L-1时出现峰值(257.97 U·g-1),此后SOD的活性迅速下降,均低于对照值239.33 U·g-1,表明连香树芽苗对水分胁迫极为敏感,在轻微的干旱胁迫下就能诱导SOD活性达到高峰,但严重胁迫则抑制了其活性。各处理间差异极显著。
2.2.6 对过氧化物酶(POD)活性的影响POD在植物体内具有广泛的作用,其主要作用之一是催化H2O2降解。与对照相比,无论是PEG溶液对连香树种子的预处理,还是干旱胁迫,POD的活性都随着PEG质量浓度的增大而明显升高。质量浓度在345 g·L-1时预处理种子的POD活性最高(133.77 U·g-1),为对照值的1.53倍,不同质量浓度的PEG引发处理均与对照差异极显著; 质量浓度221 g·L-1时干旱胁迫下POD活性最高(115.27 U·g-1),为对照值的1.66倍,不同质量浓度的PEG干旱胁迫处理均与对照差异极显著。
2.2.7 对过氧化氢酶(CAT)活性的影响种子吸胀过程中,生理活动恢复加强,不可避免地与体内生物大分子蛋白质、核酸和脂类等起反应且产生破坏其结构和功能的H2O2等自由基,CAT可把H2O2分解为H2O和O2,以清除H2O2的毒害作用。现在普遍认为植物受到环境胁迫后,保护酶活性下降。在人为的环境干旱胁迫后,CAT的活性随着PEG质量浓度的增大而明显降低(骆建霞等,2006; 喻晓丽等,2007),试验发现在干旱胁迫下不同质量浓度PEG的连香树芽苗中CAT活性均明显低于对照,质量浓度221 g·L-1时POD活性最低(10.70 mg·g-1),仅为对照值的0.48倍,各处理间差异极显著。但从试验结果来看,在PEG预处理种子后,CAT的活性随着PEG质量浓度的增大却呈上升趋势,表明PEG质量浓度过高引起种子代谢紊乱、造成无氧呼吸上升、大量消耗贮藏物质,使得CAT活性升高。不同质量浓度PEG的连香树芽苗中CAT活性均明显高于对照,质量浓度为345 g·L-1时CAT活性最高(30.10 mg·g-1),为对照值的1.92倍。不同质量浓度处理均与对照差异极显著。
3 结论与讨论PEG是一种高分子渗透剂,用其处理的实质是限制水分进入种子的速率,从而减少了种子吸胀过程中膜系统的损伤,也有利于膜系统损伤的修复。试验材料采后经过贮藏约2年,此时的种子活力下降,膜脂过氧化较为严重。试验结果表明,用不同质量浓度的PEG对连香树种子进行预处理,其MDA含量与对照相比均有不同程度的降低,这就更进一步验证了PEG预处理降低了脂质过氧化反应,有利于膜系统损伤的修复(Stewert et al., 1980)。试验研究发现,适宜质量浓度(140~301 g·L-1)的PEG预处理连香树种子,能促进种子萌发和芽苗生长,增加渗透调节物质如可溶性蛋白、可溶性糖和脯氨酸的含量,提高保护性酶SOD,POD和CAT的活性,降低MDA含量,修复膜系统损伤,提高其抗旱性。已有研究结果表明,用PEG预处理种子,能提高出苗整齐度,最终获得苗木早、齐、壮的目的(宋丽华等,2005; 程嘉翎等,2006; 王慧超等,2008),但由于本次试验只是对PEG处理初期的连香树种子萌发和芽苗生长进行了研究,认为适当质量浓度的PEG溶液(221 g·L-1)预处理能够促进连香树种子的萌发和生长,提高其抗旱性,至于PEG在后期生长过程中的幼苗形态和生理生化变化,还有待于进一步研究和探讨。
在实验室模拟干旱胁迫条件下,连香树种子萌发及其芽苗生长能够忍受PEG质量浓度为301 g·L-1的干旱胁迫环境,但在低质量浓度的PEG(95 g·L-1)胁迫下已经开始出现不适现象,种子萌发和芽苗生长都开始受到抑制,各项生理生化指标也反映出连香树对水分胁迫较为敏感,不能适应低水势的外界环境。本试验中连香树的各项生理生化指标变化趋势也与其他植物相似(孙国荣等,2003; 马武昌等,2006; 柯世省等,2007a; 2007b;时忠杰等,2007),进一步证实PEG模拟胁迫能够引起某些保护酶活性下降,减弱机体清除自由基能力,同时显著增加MDA含量,造成生理代谢紊乱,最终导致植物受到伤害(吴志华等,2004)。
曹帮华. 2005. 刺槐抗旱抗盐特性研究. 北京林业大学博士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-10022-2006023116.htm
|
程嘉翎, 段建丽, 王娜, 等. 2006. 聚乙二醇模拟水分胁迫对桑树种子萌发和生理的影响[J]. 安徽农业科学, 34(24): 6420-6422. DOI:10.3969/j.issn.0517-6611.2006.24.007 |
柯世省, 杨敏文. 2007a. 水分胁迫对云锦杜鹃抗氧化系统和脂类过氧化的影响[J]. 园艺学报, 4(5): 1217-1222. |
柯世省, 金则新. 2007b. 干旱胁迫对夏腊梅叶片脂质过氧化及抗氧化系统的影响[J]. 林业科学, 43(10): 28-33. |
傅立国. 1991. 中国植物红皮书:稀有濒危植物[M]. 北京: 科学出版社, 第一册.
|
李合生, 孙群, 赵世杰, 等. 2001. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社.
|
李合生, 孟庆伟, 夏凯. 2002. 现代植物生理学[M]. 北京: 高等教育出版社.
|
刘奕清, 陈泽雄, 杨婉晴. 2008. 高温和干旱胁迫对尾巨桉幼苗生理特性的影响[J]. 园艺学报, 35(5): 761-764. DOI:10.3321/j.issn:0513-353X.2008.05.023 |
马武昌, 王雁, 彭镇华. 2006. 车前和紫花地丁对水分胁迫的生理反应[J]. 林业科学研究, 19(5): 633-637. DOI:10.3321/j.issn:1001-1498.2006.05.018 |
骆建霞, 史燕山, 曹鸿斌, 等. 2006. 水分胁迫对蔓生紫薇和亮叶忍冬生长及生理特性的影响[J]. 园艺学报, 33(3): 657-659. DOI:10.3321/j.issn:0513-353X.2006.03.043 |
时忠杰, 杜阿朋, 胡哲森, 等. 2007. 水分胁迫对板栗幼苗叶片活性氧代谢的影响[J]. 林业科学研究, 20(5): 683-687. DOI:10.3321/j.issn:1001-1498.2007.05.016 |
宋丽华, 刘雯雯, 陈淑芬. 2005. PEG处理对臭椿种子萌发的影响[J]. 农业科学研究, 26(4): 25-29. DOI:10.3969/j.issn.1673-0747.2005.04.007 |
孙国荣, 彭永臻, 阎秀峰, 等. 2003. 干旱胁迫对白桦实生苗保护酶活性及脂质过氧化作用的影响[J]. 林业科学, 39(1): 165-167. DOI:10.3321/j.issn:1001-7488.2003.01.027 |
王慧超, 何士敏, 李昌满. 2008. PEG渗调处理对植物种子的影响[J]. 安徽农业科学, 36(6): 2224-2226. DOI:10.3969/j.issn.0517-6611.2008.06.032 |
吴志华, 曾富华, 马生健, 等. 2004. 水分胁迫下植物活性氧代谢研究进展(综述Ⅰ)[J]. 亚热带植物科学, 33(2): 77-8O. DOI:10.3969/j.issn.1009-7791.2004.02.023 |
叶功富, 陈如凯, 张水松, 等. 1999. 水分胁迫对木麻黄细胞膜稳定性和细胞保护酶影响的研究[J]. 福建林业科技, 26(增): 6-8. |
喻晓丽, 邸雪颖, 宋丽萍. 2007. 水分胁迫对火炬树幼苗生长和生理特性的影响[J]. 林业科学, 43(11): 57-61. |
张殿忠, 汪沛洪, 赵会贤. 1990. 测定小麦叶片游离脯氨酸含量的方法[J]. 植物生理学通讯, 26(4): 62-65. |
张志良. 2003. 植物生理学实验指导[M]. 北京: 高等教育出版社, 127-128.
|
赵世杰, 许长成, 邹琦, 等. 1994. 植物组织中丙二醛测定方法的改进[J]. 植物生理学通讯, 30(3): 207-210. |
Bradford M M. 1976. A rapid and sensitive method for the quantitation of micro gram quantities of protein utilizing the principle of protein-dye binding[J]. Anal Biochem, 2: 248-254. |
Christophe C, Norbert E. 1996. Detection, isolation and structure elucidation of a chlorophyll a catabolite from autumnal senescent leaves of Cercidiphyllum japonicum[J]. Phytochemistry, 42(6): 1531-1536. DOI:10.1016/0031-9422(96)00155-0 |
Dhindsa R S, Plumb-Dhindsa P, Thorpe T A. 1981. , Leaf senescence:Correlated with increased leaves of membrane permeability and lipid peroxidation and decreased levels of superoxide dismutase and catalase[J]. J Exp Bot, 32: 93-101. DOI:10.1093/jxb/32.1.93 |
Gibbons B. 1995. Tree of Britian and Europe[M]. London: Chancer Press.
|
Stewert R C, Bewley J D. 1980. Lipid peroxidation associated with accelerated aging of soybean axes[J]. Plant Physiol, 65: 245-248. DOI:10.1104/pp.65.2.245 |