 2009, Vol. 45
2009, Vol. 45文章信息
- 刘青华, 金国庆, 张蕊, 储德裕, 秦国峰, 周志春.
- Liu Qinghua, Jin Guoqing, Zhang Rui, Chu Deyu, Qin Guofeng, Zhou Zhichun
- 24年生马尾松生长、形质和木材基本密度的种源变异与种源区划
- Provenance Variation in Growth, Stem-Form and Wood Density of Masson Pine at 24-Year-Old and the Provenance Division
- 林业科学, 2009, 45(10): 55-61.
- Scientia Silvae Sinicae, 2009, 45(10): 55-61.
-
文章历史
- 收稿日期:2008-08-17
-
作者相关文章



2. 浙江省淳安县姥山林场 淳安 311700
2. Laoshan Forest Farm of Chun'an County of Zhejiang Province Chun'an 311700
木材和石油是国家的两大战略资源,但与石油不同,木材是可再生的重要自然资源,广泛用-于燃料、建筑、造纸和家具等。对森林进行有效的遗传调控和高效栽培,木材产量可成倍地-增长。木材是一种变异性很大的物质,树种之间、种内不同地理种源间、种源内不同个体之-间以及个体内皆存在着差异(Bamber et al., 1983;Megraw,1985)。当-将一个树种引种至新环境,其木材性质会发生较大的变化,有时难以预测。为改良木材和最-终产品性质及有效利用木材,必须认识和研究材性的变异模式、所受遗传控制、生长和环境-影响及木材性质对产品的影响等。
种源试验和优良种源选择是林木遗传改良最基础的工作。1821年法国De Vilmorin最早开展了林木种源试验,之后欧美等国相继系统开展欧洲赤松(Pinus sylvestris)、挪威云杉(Picea abies)和火炬松(Pinus taeda)等种源试验,证实了来自不同产地的种源其生长、材性和适应性等存在显著的遗传差-异,并充分利用这种遗传变异(Blumenrother et al., 2001)。如Shutyaev等(2000)研究发现来自欧洲西南部和东欧平原的欧洲赤松种源生产力水平较高,并依据多个区域试验点的种源试验结果将欧洲赤松划分为10个种源区。火炬松的种源试验发现,来自美国东海岸和墨西哥湾的种源生长迅速,佛吉尼亚及马里兰州的种源生长缓慢但耐寒,密西西比河以西的种源抗锈病和耐干旱(潘志刚,1992)。火炬松西部和东北部的种源木材密度较高,而南部和东南部种源的木材密度较低(Tauer et al., 1990)。我国主要造林树种的种源试验始于20世纪70年代中后期,已基本揭示了杉木(Cunnighamia lanceolata)(洪菊生等,1994)、马尾松(Pinus massoniana)(全国马尾松地理种源试验协作组,1987)、油松(Pinus tabulaeformis)(徐化成,1992)、落叶松(Larix)(杨书文,1994)等树种幼林期主要经济性状的种源差异和地理变异模式,分别不同造林区初选了一批优良种源供生产应用,并对这些树种进行种子区划以指导种子的科学调拨(徐化成,1990)。
马尾松是我国松属中分布最广的树种,生长快,适应性强,与杉木并列为南方山地的两大针叶商品用材树种,广泛用于制浆造纸、建筑、松香等。1981年和1984年由中国林业科学研究院亚热带林业研究所组织开展了2次大规模的马尾松全分布区种源多点区域试验,已揭示了苗期和幼林生长、形态等种源变异和地理变异模式,并将马尾松划分为北带、中带、南带和四川4个种源区和4个种源亚区(全国马尾松地理种源试验协作组,1987)。幼林试验发现,来自马尾松自然分布区南部的种源其生长量大,木材密度较小,福建西部、江西中南、湖南南端、广东西北和广西东南这个带状区域是马尾松造纸材的优良种源区(周志春等,1993)。秦国峰等(1995)还通过制浆造纸试验选出了6个马尾松纸浆材最佳种源。然而上述皆是幼林阶段的试验结果,无法说明马尾松成材时生长和材性的种源变异规律和种源选择结果。鉴于此,本文利用设置在浙江省淳安县姥山林场保存完好的24年生49个产地的马尾松种源试验林,研究已达主伐年龄时马尾松生长、形质和木材基本密度的种源差异和地理变异模式,通过聚类分析深入开展马尾松种源区划分,并分别纸浆材和锯材培育目标筛选一批速生优质的马尾松优良种源。
1 材料与方法 1.1 材料来源试验材料取自设置在浙江省淳安县姥山林场的24年生马尾松种源试验林,计有来自14个省区49个产地的种源参试(图 2)。该试验林位于千岛湖的东南湖区(119°02′55″E,29°33′30″N),年均温17.0 ℃,年降水量1 430 mm,无霜期263天,年日照时数1 951 h,海拔150 m,坡度20~30°,土壤为山地黄壤,肥力中等,土层厚度在80 cm以上。试验采用完全随机区组设计,8次重复,8株双列小区,株行距2 m×2 m,块状整地,穴规40 cm×40 cm×30 cm。试验林于1984年春利用1年生裸根苗造林,造林成活率在95%以上。造林后连续抚育3年至幼林郁闭。分别在造林后第10年和第15年进行2次间伐,伐去试验小区中的被压木,每小区保留生长正常的植株3~4株。

|
图 2 参试种源的Q型聚类分析树型图 Figure 2 Dendrogram of hierarchical cluster analysis for tested provenances AH: Anhui; CQ: Chongqing; FJ: Fujian; GD: Guangdong; GX:Guangxi; GZ: Guizhou; HN1: Henan; HN: Hunan; HB: Hubei; ZJ: Zhejiang; JX: Jiangxi; JS: Jiangsu; SC: Sichuan; SX: Shaanxi. |
由于重复Ⅰ和Ⅳ保存不完整,这里只利用其余6个重复的材料。2008年4月在每个重复的试验小区中选择2株生长最佳植株,测量树高、胸径、树干通直度、树干圆满度(用树干下部3 m高处直径与胸径之比表示)等指标。树干通直度按通直、较通直、一般、弯曲、严重弯曲5级,分别记分为5,4,3,2和1,分数越高越通直。同时在植株胸高上坡方位用6 mm直径的生长锥钻取一髓心至树皮的完整无疵木芯。对所取木芯自髓心向外,每5个年轮切成一段,量测其宽度(Wi),并用最大含水量法(Smith,1954)测定每年轮段的木材基本密度(Di)。根据每个重复的立地条件,将6个重复构建成一个从较差→一般→较好→很好的一维环境梯度,可用不同环境梯度的测定值简单地评价微立地条件对各性状的影响。
1.3 数据分析利用各年轮段宽度测定值估算其所代表的年轮段面积(Ai),进而计算整个木芯的加权平均基本密度Db=∑AiDi/∑Ai。单株材积(V)按公式V=0.000 062 341 803×D1.31.856 149 7× H0.956 849 2估算(周志春等,1993)。以小区单株测定值为单元,采用SAS/STAT软件中的GLM程序进行性状方差分析,以验证重复、种源和种源×重复互作效应。性状遗传相关分析时则以小区平均值为单元。方差分析和遗传相关分析时树干通直度经 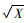
方差分析结果(表 1)显示,马尾松生长、形质和木材基本密度等主要经济性状皆存在显著或极显著的种源效应。24年生时种源胸径变幅为13.20~26.88 cm,平均值为20.14 cm,胸径生长量最大种源是最小种源的2.04倍,如广西岑溪、恭城、福建永定、大田等种源平均胸径皆在25 cm以上,安徽霍山和湖北红安2种源的平均胸径分别为13.90 cm和13.20 cm,仅为上述速生种源的50%左右。相对于种源胸径生长,种源间单株材积生长变异最大,变幅范围为0.103 1~0.462 4 m3,材积生长量最大种源是最小种源的4.48倍。种源间树高生长差异相对较小,变异系数为13.25%,仅为种源胸径变异系数的56.10%。
|
|
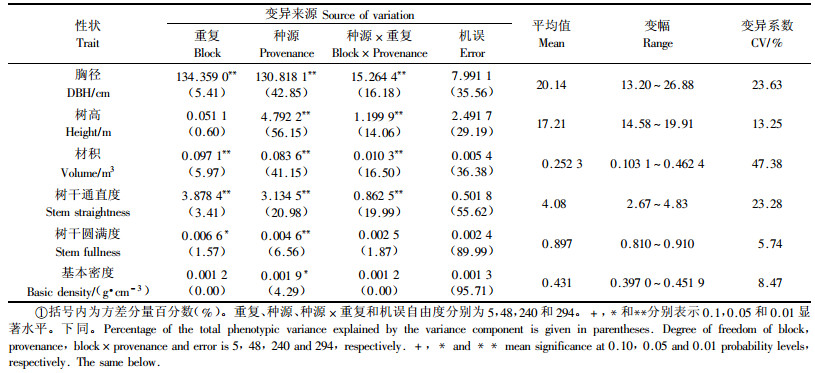
马尾松不同种源的树干通直度差异显著,如来自广东、广西、贵州、湖南、福建等速生种源树干通直,得分均在4分以上,而湖北红安、陕西城固慢生种源的干形较差,得分仅为2.67和2.90。相对于树干通直度,马尾松树干圆满度和木材基本密度的种源差异相对较小,种源树干圆满度变幅为0.810~0.910,变异系数为5.74%;种源木材基本密度变幅为0.397 0~0.451 9 g·cm-3,最大和最小种源相差13.8%。从表 1可以看出,树干圆满度和木材基本密度的种源变异仅分别占总变异的6.56%和4.29%,较多的变异来自种源内个体间,因此在充分利用种源变异的同时,还应重视种源内个体的选择。
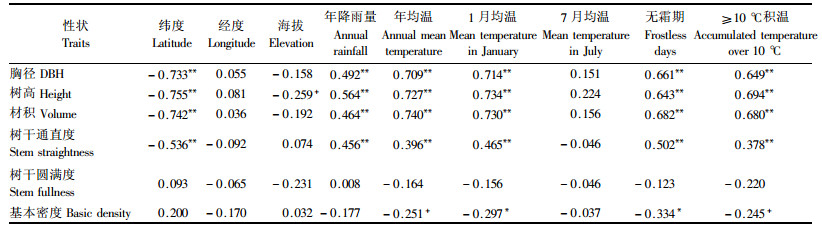
2.2 生长、形质和木材基本密度地理变异模式及与产地气候因子关系由表 2知,24年生的马尾松种源胸径、树高和单株材积生长与产地纬度呈显著的负相关,与产地经度的相关性则较小。因参试的49个种源其原产地海拔在8.9~835 m,其中近90%的种源其产地海拔低于600 m,因此海拔效应较小,仅种源树高与产地海拔的负相关达10%显著性概率水平。马尾松种源胸径、树高和材积生长主要呈典型的纬向倾群变异模式,这种纬向变异模式主要是由原产地的水(年降雨量)热(年均温、1月均温、无霜期、≥10 ℃积温)资源条件差异引起的。来自低纬度地区的种源其原产地年降雨量大、积温高、生长期长,因此种源生长量大;随着纬度升高,水热资源条件变差,尤其是产地温度成为生长的主要限制因子,因此来自北部地区的种源树高、胸径和材积生长量明显地低于南部种源。马尾松种源树干通直度也呈典型的纬向地理变异模式,这也与原产地的年降雨量、年均温、1月均温、≥10 ℃积温和无霜期有关,南部种源的树干通直度明显地高于北部种源。种源树干圆满度与产地经纬度不相关,表现为随机的地理变异模式。种源木材基本密度与产地经纬度相关性也较小,分别呈弱度的经向正相关和弱度的纬向负相关,但却与产地的年均温、1月均温、无霜期和≥10 ℃积温呈显著的负相关,即来自产地年均温和1月均温高、无霜期长、积温高的种源木材基本密度较低。
|
|
由表 1可知,除木材基本密度和树高外,其他生长和形质性状的微立地环境(重复)效应都达到1%或5%的显著水平。图 1显示,随着微立地条件的改善,马尾松胸径和材积生长量明显增加,在很好立地上24年生时的种源平均胸径和单株材积分别为21.18 cm和0.286 2 m3,是较差立地上1.17倍和1.56倍。在立地环境梯度上种源树干通直度差异显著,在很好和较好立地上种源树干通直度平均得分分别为4.07和4.27,而在较差和一般立地上分别得分3.77和4.00。种源树干圆满度也随微立地条件的改善表现出递增趋势,但增幅相对较小,从较差立地时的0.883增至很好立地时的0.903。

|
图 1 微立地环境梯度上种源性状均值 Figure 1 Trait mean over a microsite environment gradient |
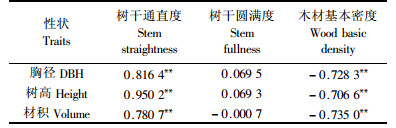
性状相关分析发现,24年生时的马尾松种源胸径、树高和单株材积与树干通直度呈显著的正遗传相关(rG=0.780 7~0.816 4),而与木材基本密度则呈显著的负遗传相关(rG=-0.706 6~-0.735 0),意味着在选择生长快的速生种源时可同时改良其树干通直度,但木材基本密度却因此会明显降低。在种源水平上,马尾松树高、胸径和材积生长对树干圆满度的影响很弱,在遗传上互为独立,选择速生种源不会导致树干的尖削度变大。
|
|
采用生长、形质和木材基本密度等6个性状的种源均值,对参试种源进行Q型聚类(图 2)。从图中可以看出,若以欧氏距离10为阈值,可将49个种源明显地划分为3个大组也即南部、中部和北部3个种源区。南部种源区(图 3中★表示)范围包括两广、福建、湖南南部、江西南部和浙江南部区域,该种源区的种源主要分布于武夷山脉、南岭山脉和云开大山地,其特性是生长量大,树干通直,24年生种源平均胸径、树高和单株材积分别为24.00 cm、18.66 m和0.362 1 m3,通直度平均得分4.42。中部种源区(图 3中●表示)包括贵州、湖南中北部、江西中北部、湖北南部、安徽南端、四川中东部、浙江中北部,也即我国亚热带中部和中北部的广大地区,该种源区的种源生长量中等,树干通直度较好,其种源平均胸径、树高和单株材积分别为19.08 cm,16.78 m和0.216 6 m3,通直度平均得分4.10。广东博罗和福建南靖两种源在地域上虽处于南部种源区范围内,但因两者皆位于马尾松自然分布区南部的边缘,生长表现略差而与中部种源聚为一类。马尾松自然分布区的北缘即为北部种源区(图 3中▲表示),主要包括大别山、大巴山以及川西种源,该种源区的种源生长较差,树干通直度一般,种源平均胸径、树高和单株材积分别为15.18 cm,15.59 m和0.136 1 m3,通直度平均得分仅3.36。福建仙游种源地处福建东部沿海地区,生长也较差,与北部种源聚为一类。相对于南部种源区的种源,中部和北部种源区的种源欧氏距离较近。

|
图 3 马尾松种源区划分 Figure 3 Allocation of provenance zones of P. massoniana |
马尾松主要用作纸浆材和锯材,2种用材对材质材性要求不同。纸浆材选育目标要求速生丰产,单位时间单位面积的纸浆材收获量最大。这里以单株木材干物质产量(单株材积与木材基本密度的乘积)为选择指标,以当地浙江淳安种源为对照,筛选出19个马尾松纸浆材优良种源(表 4),包括广西岑溪、横县、恭城,福建永定,广东焦岭等17个南部种源区的种源及2个中部种源区的种源,24年生时平均单株木材干物质量在114.81~184.84 kg,较当地浙江淳安种源高出20.9%~94.6%。
|
|
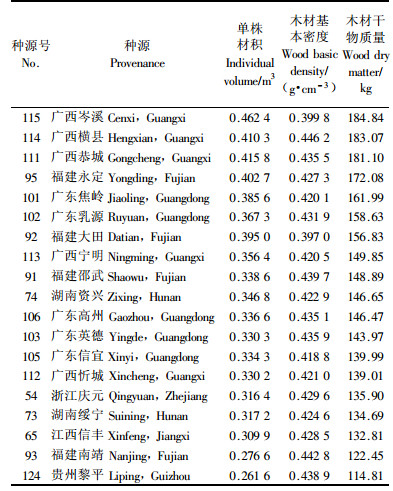
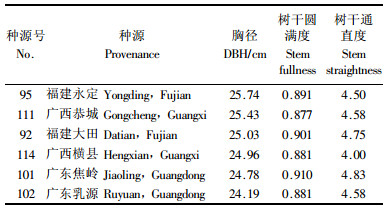
锯材优良种源要求径生长量大、木材密度高、干形好。以胸径大于浙江淳安种源20%,木材基本密度、树干通直度和圆满度形质指标不低于或不差于浙江淳安种源作为约束条件,共选出福建永定、大田,广西恭城、横县,广东焦岭和乳源6个锯材优良种源(表 5)。
|
|
本文利用已达主伐年龄的马尾松全分布地理种源试验,发现24年生的马尾松种源胸径、树高、单株材积、树干通直度等在种源间差异显著,与产地纬度呈显著的负相关,来自南部和偏南部的种源生产力水平高且树干通直,进一步证实了马尾松种源生长等性状呈明显的纬向倾群地理变异模式(全国马尾松地理种源试验协作组,1987; 周志春等,1993),这种变异模式是由其产地水热资源条件差异引起的。马尾松天然林木材密度与产地纬度呈显著负相关,自然分布区南部天然林分的木材密度显著高于北部的天然林分(周志春等,1995)。然而试验研究发现,24年生时马尾松种源木材基本密度与产地经度和纬度分别呈弱度的负相关和正相关,地理变异模式不明显,但种源木材基本密度却与产地1月均温、无霜期和≥10 ℃积温呈显著负相关,即来自气温低、无霜期短的产地的种源木材基本密度较高,这有异于周志春等(1993)和徐立安等(1997)对10~12年生马尾松种源试验的研究结果。周志春等(1993)发现10年生时马尾松种源木材基本密度随纬度的增高而增大,来自分布区西北的种源木材基本密度最大。徐立安等(1997)研究结果认为12年生时马尾松种源木材基本密度表现出西南—东北走向,东部偏北的种源木材密度较大,西南部种源的基本密度则较小。造成这种差异的可能原因是研究结果受性状年龄和试验地点的影响。为此,在该试验的基础上,将利用不同年轮段的种源木芯样品,研究不同种源木材基本密度的径向变异规律,找出造成不同年龄、不同试验点马尾松木材基本密度不同变异规律的影响因素。
了解生长与材性的遗传相关有助于速生优质林木新品种的选育。Zobel等(1989)认为多数针叶树尤其是硬松类树种其生长与木材密度相关性很小。本文研究发现,24年生马尾松种源胸径、树高和单株材积与树干通直度呈显著的正遗传相关,而与木材基本密度呈显著的负遗传相关,进一步证实了马尾松已有研究结果,即选择生产力水平高的速生种源其木材基本密度会明显降低(周志春等,1990;1993)。与树干通直度不同,生长对树干圆满度的影响较小。考虑到胸径、树高、材积和树干通直度具有较大的种源变异,而木材基本密度的遗传变异则较多地来源于种源内个体间,因此应坚持的选择育种策略为:在种源水平,应以生长量和树干通直度改良为主,材性改良为辅;而在种源内个体水平上,可依据培育目标的不同,赋予生长和木材基本密度等性状以不同的权重进行联合选择。
本文根据24年生时的生长、形质和木材基本密度将马尾松划分为南部、中部和北部3个种源区,而全国马尾松地理种源试验协作组(1987)利用幼龄林种源试验结果却将马尾松划分为北带、中带、南带和四川4个种源区,南带和中带种源区又分别划分为东西2个种源亚区。2种划分存在一定的差异。这与不同种源随时间的生长发育特性差异有关,如闽北、闽西(如福建邵武、大田)和浙南(浙江庆元)的种源早期生长速度不及两广种源,但后期生长却很强劲,本试验将其归为南部种源区,而幼龄时却将其归为中带种源区(东区亚区)。幼龄时单独区划1个四川种源区,本文研究发现,四川东部种源生产力水平中等而划为中部种源区,四川西部种源生长较慢、干形较差而归入北部种源区。浙江北部的种源后期生长也较好,本文也将其归入中部种源区。由于南部和中部种源区内的种源差异较小,本文未进一步划分为东部和西部种源亚区。本试验的种源区划分结果也充分说明了长期种源试验的重要性。这里以纸浆材和锯材为培育目标,分别选出19个和6个优良种源,其来源区域大致与周志春等(1993)确定的马尾松造纸材优良种源区相同,但范围有所扩大,包括了闽北和浙南的种源。
洪菊生, 陈延新. 1994. 全国杉木种源试验专刊[J]. 林业科学研究, 7(专刊): 1-15. |
潘志刚. 1992. 火炬松、湿地松种源试验研究[M]. 北京: 北京科学技术出版社.
|
秦国峰, 周志春, 李光荣, 等. 1995. 马尾松造纸材最优产地的确定[J]. 林业科学研究, 8(3): 266-271. DOI:10.3321/j.issn:1001-1498.1995.03.003 |
全国马尾松地理种源试验协作组. 1987. 马尾松种源变异及种源区划分的研究[J]. 亚热带林业科技, 15(2): 81-89. |
徐化成. 1990. 林木种子区划[M]. 北京: 中国林业出版社.
|
徐化成. 1992. 油松地理变异和种源区划[M]. 北京: 中国林业出版社.
|
徐立安, 陈天华, 王章荣, 等. 1997. 马尾松种源子代材性变异与制浆造纸材优良种源选择[J]. 南京林业大学学报, 21(2): 1-6. |
杨书文. 1994. 落叶松的遗传改良[M]. 哈尔滨: 东北林业大学出版社.
|
周志春, 傅玉狮, 吴天林. 1993. 马尾松生长和材性的地理遗传变异及最优种源区的划定[J]. 林业科学研究, 6(5): 556-564. DOI:10.3321/j.issn:1001-1498.1993.05.001 |
周志春, 金国庆, 秦国峰. 1990. 马尾松幼龄材密度、管胞长度的地理遗传变异及性状相关[J]. 林业科学研究, 3(4): 393-397. |
周志春, 秦国峰. 1995. 马尾松天然林木材化学组分和浆纸性能的地理模式[J]. 林业科学研究, 8(1): 1-6. DOI:10.3321/j.issn:1001-1498.1995.01.001 |
Bamber P K, Burley J. 1983. The wood properties of radiate pine. Slough, England: Commonwealth Agricultural Bureau, 1-84.
|
Blumenrother M, Bachmann M, Muller-Starck G. 2001. Genetic characters and diameter growth of provenances of Scots Pine(Pinus sylvestris L.)[J]. Silvae Genetica, 50(5/6): 212-222. |
Megraw R A. 1985. Wood quality factors in loblolly pine[J]. Georgia: TAPPI Press Atlanta, 68(7): 1-88. |
Shutyaev A M, Giertych M. 2000. Genetic subdivision of the range of Scots pine(Pinus sylvestris L.) based on a transcontinental provenance experiment[J]. Silvae Genetica, 49(3): 137-151. |
Smith D M. 1954. Maximum moisture content method for determining specific gravity of small wood samples. US Forest Service, Forest Products Laboratory, Report No: 2014.
|
Tauer C G, Loo-Dinkins J A. 1990. Seed source variation in specific gravity of loblolly pine grown in a common environment in Arkansas[J]. Forest Science, 36(4): 1133-1145. |
Zobel B J, van Buijtenen J P. 1989. Wood variation: its causes and control[M]. Berlin: Springer-Verlag, 157-173.
|