 2009, Vol. 45
2009, Vol. 45文章信息
- 郝朝运, 张小平, 张昱, 李文良.
- Hao Chaoyun, Zhang Xiaoping, Zhang Yu, Li Wenliang
- 濒危植物永瓣藤的种群生命表与动态分析
- The Life Table and Dynamics of Populations of Monimopetalum chinense, an Endangered Plant
- 林业科学, 2009, 45(9): 79-84.
- Scientia Silvae Sinicae, 2009, 45(9): 79-84.
-
文章历史
- 收稿日期:2007-11-26
-
作者相关文章



永瓣藤(Monimopetalum chinense)为卫矛科(Celastraceae)的单属种植物,半常绿木质藤本。秦仁昌于1926年在安徽祁门县棕里村发现该种,当时拟名为祁藤,后Rehder根据其花瓣永不凋落的特点定名为永瓣藤(成文法等,1998)。目前仅少量分布于安徽南部、江西北部和湖北东南部海拔200~900 m的山坡或山谷水沟边等生境中(任秀芳等, 1992; 陈灵芝, 1999; 谢国文等, 1998),为我国特有,已被列为国家二级重点保护对象(傅立国, 1992)。永瓣藤为本地起源的单型特有属和珍稀濒危物种,其研究对揭示卫矛科植物的起源进化和相互关系,以及阐明华东植物区系的起源等均具有一定意义。
自20世纪80年代以来,学者们从地理分布、群落特征、解剖结构等方面对永瓣藤展开了一定的工作(陈灵芝, 1999; 任秀芳等, 1992),但有关其种群生态学方面的研究较少(谢国文等, 1998a; 廖军等, 2000)。以往的研究和调查显示,永瓣藤分布的片断化现象比较严重,即使在各分布地也常呈岛屿状,形成不同的群落种群。研究不同群落的生境因子差异对敏感的永瓣藤种群产生的影响具有一定价值。本文以棕里村作为调查地,通过对不同类型群落中永瓣藤种群的静态生命表、存活曲线、分布格局等方面的研究,阐明其种群生存状况及未来发展趋势,为其深入研究和保护措施的制定提供指导。
1 研究区概况安徽省祁门县棕里村地处安徽省南部山区,为永瓣藤的模式标本产地。地貌呈中山、低山、丘陵,山间盆地和狭窄的河谷相交织的特征。属亚热带湿润气候,气候温和,四季分明,雨量充沛,年平均气温15.2 ℃,年平均降水量2 120 mm,无霜期约270天。在该区域内,无论林缘、溪边、路旁或林内都有永瓣藤分布,分布面积及资源量之大非常少见,且分布生境的异质性大,群落类型多样,因此可以作为本研究的调查对象。杂木林主要位于山坡下部的林缘、溪边及附近,乔木层优势种不明显,灌木层比较发达,有短柄NFDB8 (Quercus brevipetiolata)、檵木(Loropetalum chinense)等;毛竹(Phyllostachys edulis)林为单一纯林,偶尔有枫香(Liquidambar formosana)、青冈(Cyclobalanopsis glauca)等杂生,灌木层和草本层比较稀疏;杉木(Cunninghamia lanceolata)林一般在海拔500 m以下,常成纯林,灌木层有少量的尖连蕊茶(Camellia cuspidata)、映山红(Rhododendron simsii)、檵木等,草本层有海金沙(Lygodium japonicum)、翠云草(Selaginella uncinata)和苔草(Carex spp.)等;马尾松(Pinus massoniana)林以马尾松为单优势种,伴生少量的山槐(Albizia macrophylla)、赤杨叶(Alniphyllum fortunei)等,林内乔木层的盖度很高,永瓣藤数量较少。
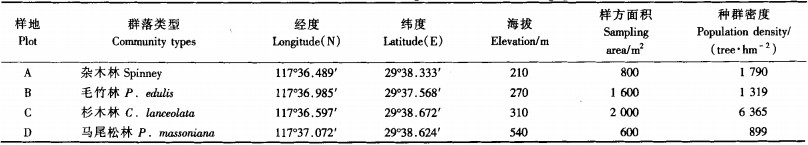
2 研究方法 2.1 取样方法及野外调查野外调查及以往研究均显示,以乔木层优势种作为划分标准,永瓣藤群落主要可分为杂木林、毛竹林、杉木林和马尾松林4种类型。采用典型样地法分别在不同类型群落中取样,结合各样地永瓣藤的分布现状及可操作性,在杂木林样地(样地A)设置2个20 m×20 m的样方,在毛竹林样地(样地B)设置一个40 m×40 m的样方,在杉木林样地(样地C)设置一个40 m×50 m的样方,在马尾松样地(样地D)设置一个20 m×30 m的样方(表 1),记录各样方的经纬度、海拔、岩石裸露率、群落透光度等生境指标。在各样方中采用相邻格子法设置5 m×5 m小样方,以小样方一角为原点,测定样方内永瓣藤个体的水平坐标(包括无性克隆分株),并用游标卡尺精确测定其基径。
|
|
小径级永瓣藤的年轮不明显(任秀芳, 1989),目前也未发现可以精确判断其生长年龄的外部特征。以往的研究认为,大小可能是比年龄更好的繁殖预测指标(Harper, 1977),许多学者在进行种群结构和动态的研究过程中都采用了大小结构分析法(Knowles et al., 1983; Johnson, 1997; 蔡飞, 2000; Rebertus et al., 1993),因此本文采用大小级结构代替年龄结构分析种群动态。
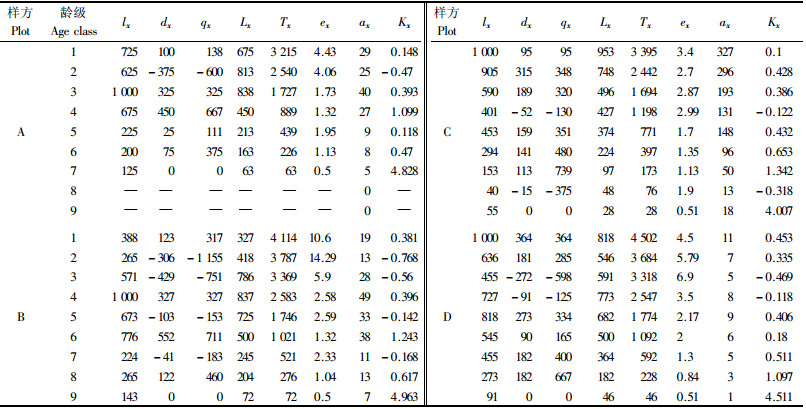
根据永瓣藤的生物学特点,同时参考其他划分方法(曲仲湘等, 1952; 刘智慧, 1990)来划分永瓣藤的径级。基径小于5 mm的永瓣藤个体分2级:根据芽鳞痕判断年龄,年龄≤2年的个体为1级,大于2年的为2级;基径≥5 mm的个体,按基径每增加2.5 mm增加1级,共分为9级。将径级从小到大的顺序看作是时间顺序关系,统计各龄级株数,参考相关文献编制种群静态生命表(陈远征等, 2006; 肖宜安等, 2004),分析其动态变化。由于各样地间的生境差异较大,因此对4个样地的永瓣藤种群分开处理。
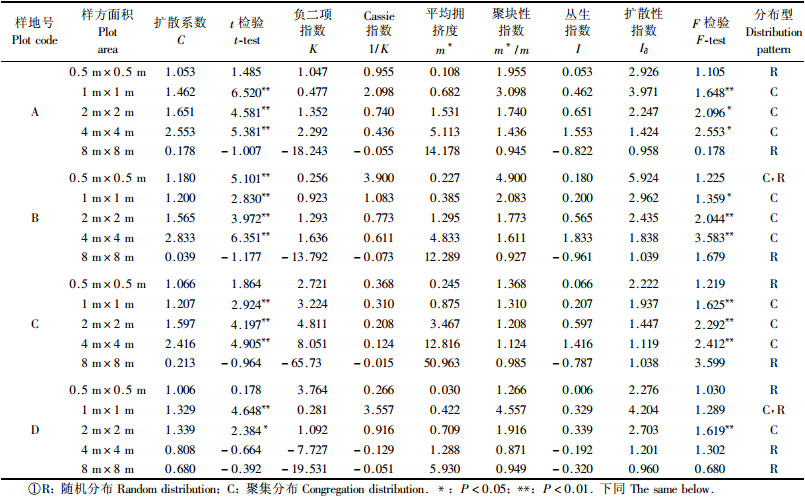
2.3 空间分布格局类型和格局强度确定以样地中永瓣藤个体定位数据为依据,分0.5 m×0.5 m,1 m×1 m,2 m×2 m,4 m×4 m、8 m×8 m共5种格子分别进行统计分析;引用扩散系数C (C值用t检验)、负二项指数(K)、平均拥挤度(m*)、聚块性指数(m*/m)、扩散型指数Iδ(Iδ值用F检验)、丛生指数(I)和Cassie指数(1/K)判定永瓣藤种群的格局类型并测定其聚集强度,分析种群不同年龄级的空间分布格局(李海涛, 1995; 郑元润, 1997; 张金屯, 2004; Greig-Smith, 1983)。
3 结果与分析 3.1 种群静态生命表及存活曲线由不同永瓣藤种群的静态生命表可知(表 2),样地A的永瓣藤个体主要集中在1,2,3,4龄级,缺失8,9龄级个体,在从4向5龄级发展过程中的消失率(Kx)极高,说明该种群缺少存活大龄永瓣藤植株的条件。群落中乔木树种较少,加之林内透光率高,永瓣藤在较低高度即可获得光照,所以个体以高度较低、胸径较小的植株为主。样地B的永瓣藤个体主要集中在3,4,5,6龄级,在幼苗时期的消失率基本为负值,说明幼苗数量不足,该种群需要补充一定数量的幼苗来维持发展,否则将走向衰退。样地C除了在7龄级的消失率较高以外,其他基本较为稳定,随着龄级的升高个体数量逐步减少,其种群状态较为理想。样地D中永瓣藤在3,4龄级的消失率均为负值,个体数量在1,5龄级存在2个峰值。整体而言,样地A,C,D中永瓣藤的幼苗数量所占比例较多,中龄和老龄个体所占比例较少,种群的更新状态较好,属于增长型或稳定型种群;样地B中个体集中在中龄阶段,幼苗所占比例较少,种群处于衰退的早期阶段。
|
|
由图 1可以看出,不同样地永瓣藤种群存活曲线存在差异。样地A中1级幼龄个体的存活率较高,3级以后死亡率显著升高,幼龄个体不足,存活曲线介于Deevey-Ⅰ型和Ⅱ型之间;B中早期个体死亡率稍高,幼龄数量稍显不足,存活曲线趋向于Deevey-Ⅰ型,种群处于衰退的早期阶段;C的存活曲线较为平滑稳定,仅在8级时死亡率较高,曲线趋向于Deevey-Ⅱ型(Manuel et al., 2002);D中永瓣藤种群的存活曲线和A的基本类似。

|
图 1 棕里村永瓣藤种群存活曲线 Figure 1 The survivorship curve of M. chinense population in Zongli village |
环境的空间异质性是影响种群空间分布格局形成和发展的主要因素之一,随着时间的变化,植物的分布格局也发生改变。按5种大小格子来分析永瓣藤种群的空间分布类型和聚集强度(表 3),结果表明:各项聚集度指标的测定结果基本一致。样地A和C在1,4和16 m2尺度下呈集群分布,对C值进行t检验均达到极显著水平(P<0.01),对Iδ进行F检验也分别达到显著水平(P<0.05)。样地B在1,4和16 m2呈集群分布,t和F检验均达到显著水平,而在0.25 m2尺度下,对C值进行t检验显示呈集群分布,对Iδ进行F检验却表明种群呈随机分布。样地D在4 m2集群分布,t检验和F检验分别达到显著和极显著水平;而在1 m2尺度时,t检验显示呈集群分布,F检验显示呈随机分布。由分析可见,永瓣藤种群的格局强度随取样尺度的变化而不同,且均呈“小→大→小”的变化趋势。但格局强度最大值和出现最大值时的面积有差异。总体而言,各样地的多个测度指标多在4 m2或16 m2时出现峰值,表明永瓣藤的聚块规模大体介于此区间。
|
|
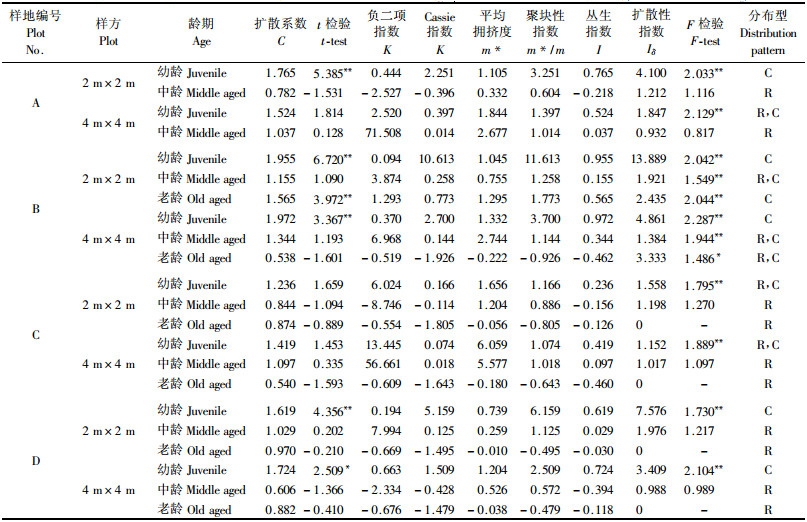
分析种群在不同发育阶段的空间格局变化可以推断种群新生个体的产生、成年个体的死亡以及种群遭受人为干扰的概况(李先琨等, 2002; 张文辉等, 2004)。将永瓣藤划分为幼龄、中龄和老龄3个阶段,分别在4~16 m2 2种尺度下分析其分布格局(表 4)。由结果可见,各龄级永瓣藤的空间分布类型有所差异,总体而言,幼龄个体多呈集群分布,中龄和老龄个体一般呈随机分布,在从幼龄→老龄的时间序列上,种群的分布格局变化趋势是从集群→轻度集群或随机。分析认为,这种扩散趋势与永瓣藤自身的生物学、生态学特性以及样地生境都有一定的关系。永瓣藤匍匐茎节上的潜伏芽在适宜的条件下萌生无性系分株苗,其幼苗基本上均是沿匍匐茎两侧呈带状分布,所以低龄阶段植株多呈集群分布,且格局规模约在4~16 m2之间。随着植株个体的发育,其对母株和外界环境的需求增加,个体间竞争加剧,物种对水分、养料、光照等资源的竞争在种内和种间产生自疏和他疏效应,种群趋于轻度集群或随机分布,与以往研究一致(祖元刚等, 1999; 郭华等, 2005)。
|
|
从研究结果可见,永瓣藤种群数量特征与其种群生物学特性及环境因素均有一定关系。杉木林内的落叶层较厚、土壤腐殖质丰富、林内透光率适中、人为干扰较少,为永瓣藤提供了良好的生存条件,匍匐茎或根系常萌发出很多无性系幼苗,种群的密度最高,更新状态较好。毛竹林内的透光率与杉木林类似,但由于林内土壤质地紧密、腐殖质含量低、持水能力差,不利于永瓣藤的无性繁殖,种群密度和稳定性均较低。马尾松林内乔木层透光率非常低,限制了永瓣藤的生存和更新,仅在一些马尾松倒伏或枯死后形成的林窗内萌发一些无性克隆苗,因此林内老龄个体所占比例较高,属于稳定型种群。次生灌丛一般是人为破坏后自然发育而来的,林内乔木层盖度低,灌木层发达,林内永瓣藤老龄个体较少,但土壤湿度大、腐殖质含量高等生境条件仍为其无性繁殖提供了一定空间。不同群落中永瓣藤种群格局强度也存在一定的差异,聚块规模大体介于4~16 m2之间。采用空间差异替代时间变化来进一步分析永瓣藤的格局动态发现,永瓣藤种群中幼龄个体大多为集群分布,在从幼龄→老龄的时间序列上,分布格局显示出扩散的趋势。
总体而言,棕里村永瓣藤种群内幼龄个体数量最多,中龄级别个体数量稍减,老龄个体最少,种群更新情况良好,这与其高效的无性繁殖有关。以往的研究显示,永瓣藤的落花落果和胚珠败育现象较普遍,导致结实率偏低;永瓣藤的种子小、种皮坚硬、种子寿命短,致使野外生境中发芽率极低甚至不发芽(谢国文, 1998)。因此,永瓣藤的有性繁殖效率极低,其种群更新主要依靠匍匐茎和根系萌生新植株,这也和野外调查结果一致。无性繁殖可以降低种群的周转、减少干扰的影响、降低种群对种子的依赖程度(Bond et al., 2001; Klimesova et al., 2004),但也存在降低种群的遗传多样性和扩散能力等不利影响,这应该是永瓣藤分布区域比较狭小的重要原因之一。同时,分析发现,永瓣藤适生于中等荫蔽、土壤含水量较高、腐殖质丰富的环境条件中,属于阴地、湿生植物,与以往研究结果一致。任秀芳(1989)对茎和叶解剖结构的研究显示,永瓣藤的木质部对水分和养料输送能力较弱;叶薄而较大,角质膜极薄,难以有效控制蒸腾;栅栏组织细胞只有一层,且体积小,海绵组织发达,叶肉细胞中的叶绿体都较大,利用光效率高;永瓣藤的根系不发达,常蔓生于表层土壤中,因此对土壤含水量的要求较高。
由于分布地距离人类活动区较近,永瓣藤种群经常遭受各种人为的干扰影响,这些干扰基本可以分为2种类型:大规模森林砍伐或垦殖人工林;具有一定频度的低强度干扰。在大规模森林砍伐或垦殖人工林的早期阶段,群落乔木层破坏所导致的林下光照强度急剧增加、土壤含水量降低、空气温度升高,将会对不耐强光和干旱的永瓣藤产生生理生态胁迫而不再适宜其生存。但是随着人工经济林或次生林的发育,群落生境将得到重建而又符合永瓣藤的生态需求,这时就会有2个可能:1)如果附近地区的永瓣藤种群均消失,那么由于有限的扩散能力永瓣藤种群很难获得重建机会,从而导致其地理分布的缩减和片断化。如原有分布记载的贵溪、宜丰、万载等县,由于解放后大规模的毁林开荒,近年来没有再次调查到永瓣藤种群的存在,很可能已经灭绝(谢国文等, 1999)。2)如果邻近群落中存有永瓣藤种群,那么其将可能再次扩散到该区域中而建立种群。因此这种干扰对永瓣藤种群的破坏将有可能随着时间的延续而得到弥补,甚至新种群的密度和规模更大。从本文结果就可以看出,分布在天然林内的永瓣藤种群一般规模较小、密度较低,反而是一些人工林或次生林内的永瓣藤种群状态更加理想。低度干扰主要是零星的薪柴砍伐、毛竹林疏伐、竹笋或药材采挖等,这种干扰行为不足以对群落结构造成重大影响,并能在一定程度增加林内的光照条件,更加有利于林下永瓣藤的生存。
蔡飞. 2000. 杭州西湖山区青冈种群结构和动态的研究[J]. 林业科学, 36(3): 67-72. DOI:10.3321/j.issn:1001-7488.2000.03.011 |
陈灵芝. 1999. 中国的生物多样性现状及其保护对策[M]. 北京: 科学出版社.
|
陈远征, 马祥庆, 冯丽贞, 等. 2006. 濒危植物沉水樟的种群生命表和谱分析[J]. 生态学报, 26(12): 4267-4272. DOI:10.3321/j.issn:1000-0933.2006.12.046 |
成文法, 姚承霞. 1998. 永瓣藤[J]. 植物杂志, (3): 3. |
傅立国. 1992. 中国植物红皮书——稀有植物(第一册)[M]. 北京: 科学出版社.
|
郭华, 王孝安, 肖娅平. 2005. 秦岭太白红杉种群空间分布格局动态及分形特征研究[J]. 应用生态学报, 16(2): 227-232. DOI:10.3321/j.issn:1001-9332.2005.02.007 |
李海涛. 1995. 植物种群分布格局研究概况[J]. 植物学通报, 12(2): 19-26. |
李先琨, 苏宗明, 向悟生, 等. 2002. 濒危植物元宝山冷杉种群结构与分布格局[J]. 生态学报, 22(12): 2246-2252. DOI:10.3321/j.issn:1000-0933.2002.12.031 |
廖军, 王宗德, 严伍明, 等. 2000. 永瓣藤植物群落种间联结研究[J]. 江西农业大学学报, 22(1): 50-53. |
刘智慧. 1990. 四川省缙云山栲树种群结构和动态的初步研究[J]. 植物生态学与地植物学学报, 14(2): 120-127. |
曲仲湘, 文振旺, 朱克贵. 1952. 南京灵谷寺森林现状的分析[J]. 植物学报, 1(1): 18-49. |
任秀芳, 蔡飞. 1992. 我国特有植物——永瓣藤花粉形态的研究[J]. 安徽师范大学学报:自然科学版, 15(3): 35-38. |
任秀芳. 1989. 我国特有植物——永瓣藤的茎和叶的解剖研究[J]. 安徽师范大学学报:自然科学版, 12(4): 22-30. |
肖宜安, 何平, 李晓红, 等. 2004. 濒危植物长柄双花木自然种群数量动态[J]. 植物生态学报, 28(2): 252-257. DOI:10.3321/j.issn:1005-264X.2004.02.017 |
谢国文, 谭策铭. 1998. 湖北新记录属植物永瓣藤种群现状及其保护[J]. 植物资源与环境, 7(4): 38-42. |
谢国文, 张志勇. 1999. 永瓣藤的地理分布及其种群空间格局[J]. 生态科学, 18(1): 7-11. DOI:10.3969/j.issn.1008-8873.1999.01.002 |
谢国文. 1998. 永瓣藤濒危因素探讨[J]. 热带亚热带植物学报, 6(1): 52-56. |
张金屯. 2004. 数量生态学[M]. 北京: 科学出版社, 243-266.
|
张文辉, 王延平, 康永祥, 等. 2004. 濒危植物太白红杉种群年龄结构及其时间序列预测分析[J]. 生物多样性, 12(3): 361-369. DOI:10.3321/j.issn:1005-0094.2004.03.009 |
郑元润. 1997. 不同方法在沙地云杉种群分布格局分析中的适用性研究[J]. 植物生态学报, 21(5): 480-484. DOI:10.3321/j.issn:1005-264X.1997.05.010 |
祖元刚, 张文辉, 阎秀峰, 等. 1999. 濒危植物裂叶沙参保护生物学[M]. 北京: 科学出版社, 182-223.
|
Bond W J, Midgley J J. 2001. Ecology of sprouting in woody plants:the persistence niche[J]. Trends in Ecology & Evolution, 16: 45-51. |
Greig-Smith P. 1983. Quantitative plant ecology[M]. Oxford: Blackwell, 68-97.
|
Harper J L. 1977. Population biology of plants[M]. London: Academic Press, 599-643.
|
Johnson J B. 1997. Stand structure and vegetation dynamics of a subalpine treed fen in Rocky Mountain National Park, Colorado[J]. Journal of Vegetation Science, 8: 337-342. DOI:10.2307/3237322 |
Klimesova J, Martinkova J. 2004. Intermediate growth forms as a model for the study of plant clonality functioning:an example with root sprouters[J]. Evolutionary Ecology, 18: 669-681. DOI:10.1007/s10682-004-5149-1 |
Knowles P, Grant M C. 1983. Age and size structure analysis of Engelmann spruce, Ponderosa pine, Loadgepole pine and Limber pine in Colorado[J]. Ecology, 64(1): 1-9. DOI:10.2307/1937322 |
Manuel C, Molles J. 2002. Ecology, concept and applications[M]. 2nd ed. New York: McGraw-Hill Companies, 186-437.
|
Rebertus A J, Veblen T T. 1993. Structure and tree-fall gap dynamics of old-growth Nothofagus forests in Tierra del Fuego, Argentina[J]. Journal of Vegetation Science, 4: 641-654. DOI:10.2307/3236129 |