 2009, Vol. 45
2009, Vol. 45文章信息
- 张茂震, 王广兴, 刘安兴.
- Zhang Maozhen, Wang Guangxing, Liu Anxing
- 基于森林资源连续清查资料估算的浙江省森林生物量及生产力
- Estimation of Forest Biomass and Net Primary Production for Zhejiang Province Based on Continuous Forest Resources Inventory
- 林业科学, 2009, 45(9): 13-17.
- Scientia Silvae Sinicae, 2009, 45(9): 13-17.
-
文章历史
- 收稿日期:2008-09-01
-
作者相关文章


2. 南伊利诺伊大学地理和环境资源系 卡本代尔 IL 62901 USA;
3. 浙江省林业调查规划院 杭州 310020
2. Department of Geography and Environmental Resources, Southern Illinois University Carbondale IL 62901, USA;
3. Zhejiang Institute of Forestry Inventory and Planning Hangzhou 310020
对于以省、流域、国家乃至全球为对象的区域森林生物量估计,方法主要有2类:基于森林资源清查数据的估计方法和基于遥感信息技术的估计方法(徐新良等,2006;郭志华等,2002;Hame et al., 1997; Dong et al., 2003; Running et al., 1991; Peterson et al., 1994)。基于森林资源清查数据的方法包括平均生物量法(Brown et al., 1984; 1989)、生物量转换因子法(Kauppi et al., 1992; Birdsey et al., 1993; Brown et al., 1992)和生物量换算因子连续函数法(variable biomass expansion factor function, variable BEFF)(Fang et al., 2001;方精云等,1996; 2002)。目前,这些方法在森林生物量/碳估算及全球碳循环研究中被广泛使用。
大尺度的区域森林生物量的估算方法一直是人们关注的焦点,自20世纪70年代以来很多学者根据国际生物学计划(IBP)时代在全球各地的实测资料,采用平均生物量的方法开始对全球范围内的生物量进行估测(Woodwell et al., 1978;Walker et al., 1997),然而由于当时各类森林的实测资料较少,而且实测生物量都普遍偏高,使得平均生物量偏大,最终导致全球和区域生物量的估算结果偏高(方精云等,1996)。与此同时,人们开始尝试通过生物量与材积的比值将森林资源蓄积转化为森林生物量。在这种简易方法中,比值被看作是一恒定的常数(Lieth et al., 1975),但实际情况并非如此,它随着材积的变化而变化,只有当材积很大时该值才逐步趋于稳定(方精云等,2001),换算因子连续函数法是材积源方法的改进。
对于区域森林生物量或森林生产力估计,大范围连续的地面调查数据极为重要。我国国家级森林资源监测体系已有30多年的历史,在这期间积累了丰富的以省为总体的森林资源连续清查数据。目前,森林资源连续清查数据除用于统计国家林业行政主管部门规定的报表以外,很少用于相关科学研究(张茂震等,2008)和进一步的决策分析。利用森林资源连续清查数据对区域森林生物量和生产力进行估计,将进一步拓展森林资源连续清查数据在科学研究和管理决策领域的应用,对区域森林碳储量估算、区域CO2源汇功能评价具有重要意义。本文利用森林资源连续清查数据和生物量换算因子连续函数法估计浙江省森林生物量和生产力,探讨大区域森林生物量和生产力估计的可行方法。
1 研究区概况浙江省位于中国东南沿海(118°01′—123°10′E,27°06′—31°11′N),地势西南部高,东北部低,自西南向东北倾斜,呈梯级下降;东北部为冲积平原,中部以丘陵为主,西南部为平均海拨800 m的山区,全省最高海拔1 929 m。全省陆域面积10.18万km2,海域面积26万km2。
浙江属亚热带季风气候,地处南亚热带和北亚热带过渡地带,日照充足,年平均气温17℃,年平均降水量1 319.7 mm;全省土壤以丘陵山地红壤、黄壤等地带性土壤和海岛饱和红壤为主,植被类型有常绿阔叶林、常绿落叶阔叶混交林、常绿落叶针叶混交林、山地矮林和山地草丛灌丛。有高等植物288科、1 471属、4 600余种,其中,木本植物107科、423属、1 407种。
根据第5次森林资源连续清查资料统计,全省林地面积667.97 hm2,活立木蓄积1.938亿m3,森林覆盖率为57.4%,林业用地面积占总面积的65.6%,幼中龄林面积占森林面积的88.2%。
2 研究方法 2.1 数据来源本研究的基础数据来自浙江省1994,1999和2004年森林资源连续清查。浙江省森林资源连续清查采用正方形样地,面积0.08 hm2。1994,1999和2004年,样地数分别为4 222,4 249和4 252个,样木数分别为121 610,149 681和207 091株。各期调查的主要技术标准基本统一。各期调查均包含的树种/树种组有柏木(Cupressus spp.)、黑松(Pinus thunbergii)、马尾松(Pinus massoniana)、黄山松(Pinus taiwanensis)、杉木(Cunninghamia lanceolata)、柳杉(Cryptomeria spp.)、栎类(Quercus spp.)、硬阔类、檫木(Sassafras tsumu)和软阔类共10个。2004年增加针叶混交林、针阔混交林、阔叶混交林3个树种组。每木检尺记录立木类型、检尺类型、树种和胸径。样地优势树种/树种组划分与树种/树种组划分标准相同。
2.2 生物量估计全省森林生物量估计采用换算因子连续函数法(Fang et al., 2001),其森林生物量估计回归方程为:B=aV+b,式中:B为单位面积生物量(t·hm-2),V为单位面积蓄积量(m3·hm-2),a和b为参数。表 1列出了12组森林蓄积量-生物量转换模型参数(Fang et al., 2001),分别对应1个或多个树种/树种组,其中,针叶混交林、针阔混交林和阔叶混交林3个树种组采用了曾伟生(2005)的参数。
|
|

在森林资源连续清查中,蓄积调查只针对经济林以外的乔木林、疏林以及散生木和四旁树进行。因此,本研究中经济林、灌木林和竹林的生物量采用非蓄积转换方法估计。其中,经济林和灌木林采用Fang等(2001)的方法估计,竹林采用周国模(2006)的方法估计。疏林、散生木及四旁树的林木生物量均由立木蓄积推算。具体计算方法和参数如下。竹林:用单株平均生物量与总株数估计,单株平均生物量取10.442 3 kg(周国模,2006)。经济林:用单位面积生物量平均值与总面积估计,单位面积生物量平均值取23.70 t·hm-2。灌木林:用单位面积生物量平均值乘以总面积估计,单位面积生物量平均值取19.76 t·hm-2(Fang et al., 2001)。疏林、散生木及四旁树:以该省同期平均单位生物量(t·hm-2)与单位蓄积量(m3·hm-2)比值(t·m-3)推算相应类型的生物量。
2.3 生产力估计森林生产力的估算方法可分为遥感和实测2类。前者基于森林光合作用机制进行模型模拟,后者基于森林某一时刻实际生物量的测定。本文采用换算因子连续函数法估算,属于后者。在已知森林蓄积的情况下:
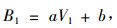
|
(1) |
式中:B1为第n1年单位面积生物量(t·hm-2),V1为第n1年单位面积蓄积量(m3·hm-2),a, b为参数。
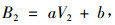
|
(2) |
式中:B2为第n2年单位面积生物量(t·hm-2),V2为第n2年单位面积蓄积量(m3·hm-2),a, b为参数。第n1年和第n2年单位面积生物量之差即为间隔期内单位面积的总生产力,令Δn=n2-n1,ΔB为单位面积单位时间生物量的增长量,则有:
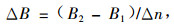
|
(3) |
即ΔB为间隔期内年均单位面积生产力。由(3)式可以导出:
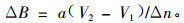
|
(4) |
公式(3)和(4)的ΔB是一个Δn年的平均值。当Δn不是太大时,2个公式均可以较好地反映森林生产力,但2式有一定区别。公式(3)用2期生物量之差计算,而(4)式直接用蓄积量之差计算,后者没用到参数b。
在总体没有人为干扰的情况下,可用上述方法计算生产力。但在大范围内,干扰不可避免。利用森林资源连续清查数据估算森林生产力,森林的采伐和枯损的因素必须考虑。因此,公式(3)和(4)分别有修正式:
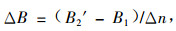
|
(5) |
式中:B′2为B2的修正值,且B′2=a[V2+(Vc+Vd)(1+p)Δm]+b, 其中Δm为采伐木和枯损木从最后1次调查到被采伐或枯损时所经历的时间(年)。取Δm=Δn/2,Vc,Vd分别为间隔期Δn年内采伐和枯损蓄积,p为年平均生长率。[(Vc+Vd)(1+p)Δm]为Δn年内的包括未测生长量(按Δm年计算)的消耗量。将B′2代入(5)式,有:

|
(6) |
本研究森林生产力计算采用修正式(5)和(6)。
3 结果与分析 3.1 生物量根据森林生物量换算因子连续函数法的回归方程和表 1,浙江省林分生物量计算结果如表 2。表 2显示, 1994, 1999和2004年浙江省林分生物量分别为0.951亿,1.006亿和1.511亿t,林分单位面积平均生物量分别为27.59,27.83和38.40 t·hm-2。1994—1999年间,林分生物量年均增长3.18%,林分面积年均递增1.07%;1999—2004年间,林分总生物量年均增长27.95%,林分面积年均递增1.68%,面积年均增长速度略有提高,但生物量增幅较大。从林分生物量与林分面积增长的速率可看出:1994—1999年间森林面积增长以幼龄林和中龄林为主,林分质量较低;1999—2004年间,林分生物量年均增长速度大大超过林分面积增长速度,表明林分质量得到大幅度改善。
|
|

林分生物量(表 2)与经济林、竹林、灌木林、疏林、散生木和四旁树生物量共同构成全省森林总生物量(表 3)。
|
|
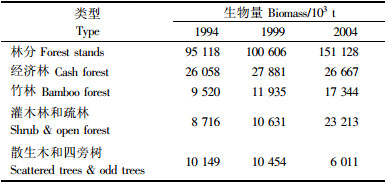
由表 2和表 3可知,浙江省1994,1999和2004年森林总生物量分别为1.496亿,1.615亿和2.244亿t,平均单位面积森林生物量分别为14.69,15.87和22.04 t·hm-2。森林总生物量变化呈上升趋势,与林分生物量变化趋势相同。1994—1999年间,森林生物量年均增长1.60%,而在1999—2004年间,这一增长率达7.78%,说明全省森林生物量在20世纪90年代稳步增长,进入21世纪以后这一增长加速,林分质量稳步提高,灌木林得到较好的保护,竹林等森林资源成分的消耗相对减少。但与其他省比较(曾伟生,2005;光增云,2006),无论是全省森林总平均单位面积生物量还是林分平均单位面积生物量都显著偏低。
3.2 生产力根据公式(5)可得,1994—1999年间全省林分平均生产力为1.557 t·hm-2a-1,其中针叶林为1.426 t·hm-2a-1,阔叶林为2.367 t·hm-2a-1。1999—2004年间全省林分平均生产力为2.060 t·hm-2a-1,其中针叶林为1.844 t·hm-2a-1,阔叶林为2.280 t·hm-2a-1。分别用公式(5)和(6)计算的各优势树种/树种组平均生产力见表 4。
|
|
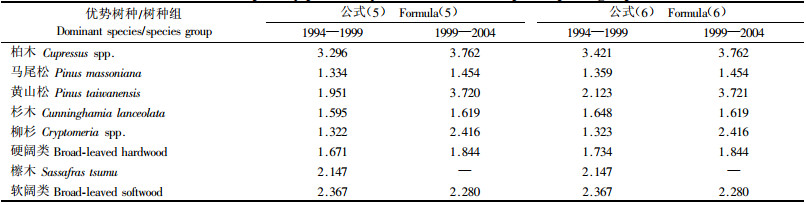
结果显示:公式(5)和(6)的计算结果基本一致。1994—1999年间平均单位面积林分生产力与1999—2004年间平均单位面积林分生产力差别明显,总体上呈上升趋势。浙江省1994—2004年间森林生产力有了显著提升,但与其他省份(曾伟生,2005;光增云,2006)相比,单位面积生物量和生产力2项指标仍显著偏低。
4 结论与讨论与全国林分生物量(方精云等,1996)比较,1994年浙江省林分生物量占1.71%,而同期活立木蓄积浙江省占全国的1.33%,二者吻合较好,说明用换算因子连续函数法估计区域森林生物量具有较高的可信度。换算因子连续函数法是估计大尺度森林生物量和生产力的有效方法,但是,采用其对单位蓄积量很低的总体进行估计,误差可能较大。
基于森林资源连续清查的区域生产力估计有生物量差函数和蓄积量差函数2种方法,前者形如ΔB=(B2-B1)/Δn,后者形如ΔB=a(V2-V1)/Δn。虽原理相同,但估计结果有一定差别。当2期林分变化较大或出现极端情况时,2种方法的结果差别显著。这里,计算B2或V2需要考虑现有森林存量和2次调查之间的采伐利用或自然损失。在本研究中,由于后期有1/3的样地移动位置,其间的采伐等消耗情况无法得知,以至于生产力有可能被低估。而后式是根据单位蓄积变化量推算生物量,不确定因素较前者少,但其未用到模型的b参数,估计结果也可能会偏低。结果证明,在基础数据分布正常的情况下,b参数的影响程度不大,采用ΔB=a(V2-V1)/Δn估算的森林生产力结果较前者更可靠。
用换算因子连续函数法估算生产力必然涉及2次调查之间的未测消耗量,如果直接采用森林资源连续清查成果提供的生长率,则由于其采用普雷斯勒式计算,计算未测消耗量不能直接用复利公式进行。另外,森林资源连续清查中的未测生长率只代表了相邻2次调查之间林木生长率的一半,而计算生产力时必须用实际生长率。因此,在实际应用中,只要数据源允许,应直接在复位样木的基础上用复利公式计算生长率,并在此基础上进行其他量和率的计算,以保证数据的一致性和准确性。
换算因子连续函数法是在森林资源连续清查基础上估算区域生物量的一种方法。据此估算的浙江省森林生物量总体上与实际情况相符,但结果的可靠性及精度仍有待于在下一步工作中通过建立本省森林生物量模型来评价。
作为大尺度森林生物量估计的一种方法,换算因子连续函数法仍有需要完善的地方,如该方法不能提供估计结果的可靠性与估计精度。关于这些问题,将通过建立浙江省森林生物量模型来进一步研究。
方精云, 刘国华, 徐嵩龄. 1996. 我国森林植被生物量和生产力[J]. 生态学报, 16(4): 497-508. |
方精云, 陈安平, 赵淑清, 等. 2002. 中国森林生物量的估算:对Fang等Science一文(Science, 2001, 291: 2320~2322)的若干说明[J]. 植物生态学报, 26(2): 243-249. DOI:10.3321/j.issn:1005-264X.2002.02.018 |
光增云. 2006. 河南森林生物量与生产力研究[J]. 河南农业大学学报, 40(5): 493-497. DOI:10.3969/j.issn.1000-2340.2006.05.010 |
郭志华, 彭少麟, 王伯荪. 2002. 利用TM数据提取粤西地区的森林生物量[J]. 生态学报, 22(11): 1832-1839. DOI:10.3321/j.issn:1000-0933.2002.11.006 |
徐新良, 曹明奎. 2006. 森林生物量遥感估算与应用分析[J]. 地球信息科学, 8(4): 122-128. DOI:10.3969/j.issn.1560-8999.2006.04.024 |
曾伟生. 2005. 云南森林生物量与生产力研究[J]. 中南林业调查规划, 24(4): 1-13. DOI:10.3969/j.issn.1003-6075.2005.04.001 |
张茂震, 王广兴. 2008. 浙江省森林生物量动态[J]. 生态学报, 28(11): 5665-5674. DOI:10.3321/j.issn:1000-0933.2008.11.052 |
周国模. 2006.毛竹林生态系统中碳储量、固定及其分配与分布的研究.浙江大学博士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-10335-2006085249.htm
|
Birdsey R A, Plantinga A G, Heath L S. 1993. Past and prospective carbon storage in United States forests[J]. For Ecol Man, 59: 33-40. |
Brown S, Gillespie A J R, Lugo A E. 1989. Biomass estimation methods for tropical forests with application to forest inventory data[J]. Forest Science, 35: 881-902. |
Brown S, Lugo A E. 1984. Biomass of tropical forests: A new estimate based on forest volumes[J]. Science, 223: 1290-1293. DOI:10.1126/science.223.4642.1290 |
Brown S, Lugo A E. 1992. Above ground biomass estimates for tropical moist forests of Brazilian Amazon[J]. Interciencia, 17: 8-18. |
Dong J R, Kaufmann R K, Myneni R B, et al. 2003. Remote sensing estimations of boreal and temporate forest woody biomass: carbon pools, sources, and sinks[J]. Remote Sensing of Environment, 84: 393-410. DOI:10.1016/S0034-4257(02)00130-X |
Fang J Y, Chen A P, Peng C H, et al. 2001. Changes in forest biomass carbon storage in China between 1949 and 1998[J]. Science, 292: 2320-2322. DOI:10.1126/science.1058629 |
Hame T, Sallli A, Andersson K, et al. 1997. A new methodology for the estimation of biomass of conifer-dominated boreal forest using NOAA AVHRR data[J]. Int J Remote Sens, 18(15): 3211-3243. DOI:10.1080/014311697217053 |
Kauppi P E, Mielik I K, Kuusela K. 1992. Biomass and carbon budget of European forests, 1971 to 1990[J]. Science, 256(5053): 70-74. DOI:10.1126/science.256.5053.70 |
Lieth H, Whittaker R H. 1975. Primary productivity of the biosphere[M]. New York: Springer-Verlad Press, 237-263.
|
Peterson D L, Waring R H. 1994. Overview of the oregon transect ecosystem research project[J]. Ecological Applications, 4(2): 210-225. DOI:10.2307/1941927 |
Running S W, Gower S T. 1991. FOREST-BGC, A general model of forest ecosystem processes for regional applications. Ⅱ. Dynamic carbon allocation and nitrogen budgets[J]. Tree Physiology, 9: 147-160. DOI:10.1093/treephys/9.1-2.147 |
Walker B H, Steffen W L, Canadell J, et al. 1997. The terrestrial biosphere and global change: Implications for natural and managed ecosystems: a synthesis of GCTE and related research. IGBP Book Series Number 4.
|
Woodwell G M, Whittaker R H, Reiners W A, et al. 1978. The biota and the world carbon budget[J]. Science, 199: 141-146. DOI:10.1126/science.199.4325.141 |