 2009, Vol. 45
2009, Vol. 45文章信息
- 康冰, 刘世荣, 蔡道雄, 卢立华.
- Kang Bing, Liu Shirong, Cai Daoxiong, Lu Lihua
- 南亚热带杉木生态系统生物量和碳素积累及其空间分布特征
- Characteristics of Biomass, Carbon Accumulation and Its Spatial Distribution in Cunninghamia lanceolata Forest Ecosystem in Low Subtropical Area
- 林业科学, 2009, 45(8): 147-153.
- Scientia Silvae Sinicae, 2009, 45(8): 147-153.
-
文章历史
- 收稿日期:2007-12-24
-
作者相关文章



2. 中国林业科学研究院森林生态环境与保护研究所 北京 100091;
3. 中国林业科学研究院热带林业实验中心 凭祥 532600
2. Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry Beijing 100091;
3. Experiment Center of Tropical Forestry, Chinese Academy of Forestry Pingxiang 532600
碳是一切有机物的基本成分,也是构成生物体的主要元素,约占生物体干质量的一半左右,碳循环及其空间分布与生态系统的维持、发展和稳定性机制有着密切的联系(裴志永等,2003)。但自从工业革命以来,人类大量燃烧化石燃料和改变土地的利用方式等原因,导致大气中的CO2浓度不断升高(Tans,2006)。近年来,有学者研究认为,大气CO2浓度每年以1.9 mL·m-3a-1的线性速率增加(Artuso et al., 2009)。根据IPCC(2001)第3次评估报告报道,在过去20年里,大气中CO2浓度每年增加速度一直保持在0.4%。由此而引起的全球性温室效应、气候变暖等生态问题已成为全世界共同关注的焦点问题之一(Norby et al., 2006)。森林作为碳汇,在全球大气碳平衡中的作用极其显著(方精云等,2001),其为减缓温室气体提供低成本的机会,而森林不同的经营管理措施(如对森林组成、年龄和结构的控制)常常影响其碳吸存的结果(Christopher et al., 2009)。目前,人工造林作为一种新增碳汇的主要途径,以木材收获与碳吸存为双重目标,已成为国际研究热点(Onigkeit et al., 2000)。杉木(Cunninghamia lanceolata)是我国特有的优良速生针叶树种,也是我国亚热带地区最主要的用材树种之一,其分布地域广阔,人工栽培、经营历史悠久,不仅提供了大量的商品用材,而且在固碳方面发挥了明显的作用。
本文研究杉木分布极限区域生态系统碳积累与分配特征,旨在了解杉木生态系统碳贮量及其潜力,为我国森林生态系统碳平衡的估算和动态模拟提供基础数据,同时也为有效提高森林碳汇能力提供参考。
1 材料与方法 1.1 试验地概况研究地点处于广西西南边陲,位于21°57′—22°19′ N,106°39′—106°59′ E。该地区位于南亚热带季风气候区域内的西南部,与北热带北缘毗邻,属湿润半湿润气候。境内日照充足,雨量充沛,干湿季节明显(10—次年3月为干季,4—9月为湿季),光、水、热资源丰富。年均气温为20.5~21.7 ℃,极端高温40.3 ℃,极端低温-1.5 ℃;≥10 ℃年活动积温6 000~7 600 ℃。年均降雨量1 200~1 500 mm,年蒸发量1 261~1 388 mm,相对湿度80%~84%。主要地貌类型以低山丘陵为主,坡度以25°~30°为多。地带性土壤为砖红壤性红壤,深度小于1 m。该区域为杉木东南部极限分布区(刘世荣等,2005)。
2005年在中国林业科学研究院热带林业实验中心大青山试验场,选择1993年栽植的杉木人工林,林龄为13年,林分郁闭度为0.85。造林地为杉木人工林采伐迹地。按坡位上、中、下分别选择3个样区(样区编号为1,2,3),每个样区设立4块固定样地,每块样地面积为20 m×30 m。林下灌木主要有对叶榕(Ficus hispida)、柘树(Cudrania tricuspidata)、白背叶(Mallotus apelta)及软荚红豆(Ormosia semicastrata)、玉叶金花(Mussaenda pubuscens)、黑龙骨(Periploca forrestii)、八角枫(Alangium chinese)等;草本主要有蔓生秀竹(Microstegium vagans)、铁线蕨(Adiantum flabellulatum)、乌毛蕨(Blechnum orientale)、凤尾蕨(Pteris nervosa)等。
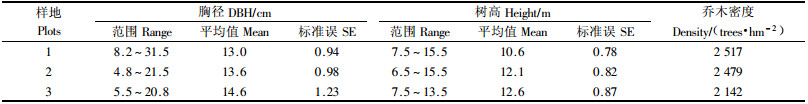
1.2 林分生物量的测定和净生产力的估算在每个固定样地,各设置6个10 m×10 m的小样方,调查各样方乔木层树高、胸径,得到乔木的组成结构(表 1)。根据固定样地的每木检尺结果,按径级(2 cm)选取标准木8株进行生物量测定。样木伐倒后,地上部分按分层切割法测定生物量,每1 m为一区分段,分层截取测定干、皮、枝、叶的生物量;地下部分采用“分层挖掘法”(0~20,20~40,40~60和60~80 cm)分别测定根头、大根、粗根、细根鲜质量。各器官抽取样品,在80 ℃烘箱中烘干至恒量,计算出各器官干物质质量(康冰等,2006)。
|
|
建立相对生长方程(如下),估算林分的生物量,以年平均增长量作为净生产力的估测指标(方晰等,2003a)。
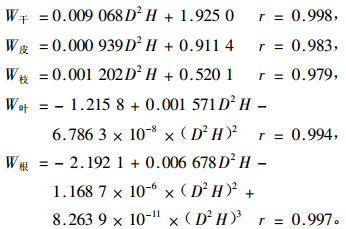
|
式中:D为胸径(cm),H为树高(m),r为相关系数。
1.3 林下植被生物量的测定在每个样地内按梅花形设置5个面积1 m×1 m的小样方,记录每个样方内的植物种类,灌木分为叶、茎、根,草本植物分为地上部分和地下部分,采用全挖法实测生物量。同种植物的相同器官取混合样品。死地被物全部测定生物量,取混合样品。将所收集的林下层植物及凋落物样品在80 ℃烘箱中烘干至恒量后,再估算干质量,同时用作分析样品。
1.4 年凋落物和林地凋落物现存量的测定在每块样地内各设置1 m×1 m的尼龙网收集器5个,每月收集凋落物1次,按组分(小枝、叶、花、果)测定干质量。同时,在每块样地内随机设置5个1 m×1 m的小样方,按未分解、半分解2个层次,测定现存凋落物的干质量。
1.5 样品采集及化学分析在测定生物量的同时,按干、皮、叶(分当年生叶和老叶)、枝(分当年生枝和老枝)、根(分 < 0.2,0.2~0.5,>0.5 cm和根头)分别采集8株标准木的分析样品,逐一进行化学分析,以样木分析结果取其平均值作为最终结果。现存凋落物是在样地的5个1 m×1 m的小样方内,先按未分解、半分解2层次分别取样,再按未分解、半分解把每个样地的5个小样方分别混合,以混合样测定作为最终结果。在不同样区内,各按0~20,20~40,40~60 cm分层随机取土样3~4次,再分层次分别混合,取混合样测定作为最终结果。同时测定其密度,按其密度估算各层单位面积土壤的质量。
植物分析样品于70 ℃烘干,土壤样品则置于室内风干,然后磨碎。所有样品均采用重铬酸钾-水合加热法测定有机碳含量(中国土壤学会农业化学专业委员会,1984)。平均值差异显著性分析用SPSS 13.0软件包进行。
1.6 碳贮量的计算方法碳贮量根据单位面积林分干物质质量(生物量)乘转换系数而求得。不同组分的转换系数是根据生物量测定时样品实测所取得的有机碳含量值。土壤碳贮量则是土壤有机碳含量、土壤密度及土壤厚度三者的乘积。土壤密度随土层加深而增加,0~20,20~40,40~60 cm土层平均值依次为1.35,1.42和1.48 g·cm-3。同一土层,各样区间土壤密度没有明显差异(P>0.05)。
2 结果与分析 2.1 植物生物量及地表凋落物现存量乔木层各器官生物量分配见图 1。杉木各器官生物量分配差异明显(P<0.05),生物量大小依次为树干>根>枝>皮>叶。

|
图 1 乔木层各器官生物量及分配 Figure 1 Biomass and its distribution in tree storey |
3个样区林分下层植物及现存凋落物量见表 2,样区1~3灌木生物量依次为1.60,0.81和0.38 t·hm-2,草本生物量依次为2.74,3.14和3.25 t·hm-2,现存凋落物量依次为7.96,7.58和9.39 t·hm-2。不论灌草,地上生物量均大于地下。林下草本生物量大于灌木生物量。林分下层植物生物量大小趋势与乔木层生物量大小相反。现存凋落物量基本与乔木生物量大小趋势一致。
|
|
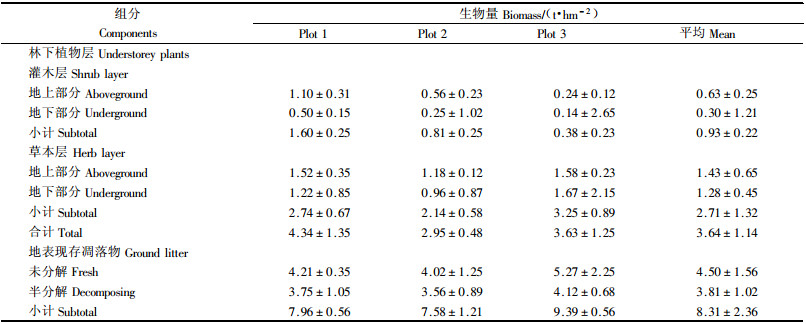
1)乔木层碳素含量 乔木层各器官碳素含量见表 3。杉木不同器官碳素含量各异,皮的含量最高,为510.5 g·kg-1,其次为叶,为501.8 g·kg-1,根和枝含量较低,变化幅度为462.1~510.5 g·kg-1。各器官碳素含量从高到低依次为:叶>皮>干>根>枝。样本的变异系数为1.85%~5.21%,各器官碳素含量差异不明显(P>0.05)。
|
|

2)林下植被及土壤碳素含量 林下植被及碳素含量的测定结果见表 4。从表 4可以看出,灌木层碳素含量平均为437.5 g·kg-1,草本层为407.8 g·kg-1,凋落物层为418.7 g·kg-1。灌木层的碳素含量高于草本层,表现出随植物个体高度或组织木质化程度的降低,其碳素含量相应减少的趋势(康冰等,2006)。不管灌木和草本,其地上部分碳素含量均高于地下部分。灌草及凋落物层碳素平均含量各组分间差异不显著(P>0.05)。
|
|

土壤各层次碳含量随着土层加深而降低,差异明显(P<0.05),变异系数为13.7%~20.2%。同一土层在各样区碳含量差异不明显(P>0.05),变异系数较小,为3.3%~8.5%(表 5)。
|
|
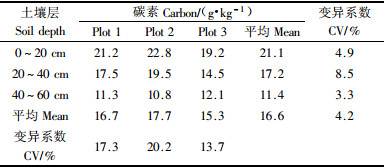
总的来看,杉木生态系统各组分碳含量有较明显的规律。在地上部分,其碳含量平均值变化规律为:乔木层>地表现存凋落物层>灌木层>草本层。在地下部分(根),表现为乔木层>灌木层>草本层。土壤的碳素含量明显低于植被层及凋落物层的碳素含量。
2.3 生态系统碳贮量及在各组分中的分配乔木层各器官碳分配特征在3个样区基本是一致的。林木各器官的生物量与相应碳素含量之积为其碳贮量,因此生物量正向影响各器官的碳贮量(表 6)。各器官中,树干的碳贮量最高,样区1~3中树干碳贮量均占乔木层碳贮量的58%以上;其次,根也占较大比例,占各自碳总贮量的19.96%~20.28%。各器官碳分配排序为:干>根>皮>枝>叶。
|
|

3个样区植被层碳贮量分别为40.43,46.31和47.54 t·hm-2(表 7),其中乔木层所占比例分别为95.09%,96.76%和96.84%。可见,植被层碳贮量主要取决于乔木层。地表现存凋落物碳贮量较小,为3.17~3.94 t·hm-2,仅是植被层的13.9%~16.89%。
|
|

样区1,2和3土壤碳贮量分别为104.39,148.94,128.48 t·hm-2。3个样区土壤碳贮量中,0~20 cm土层含碳量较高,分别占土壤层(0~60 cm)碳贮量的40.77%,41.33%和40.35%,占整个生态系统碳贮量的31.08%,31.03%和28.16%。可见,森林土壤表层碳贮量贡献较大,任何引起水土流失的活动很容易导致土壤碳损失,同时也反映了亚热带地区森林土壤的脆弱性。
杉木生态系统中碳库主要分为3个部分(表 7):植被层、凋落物层和土壤层。样区1~3中,生态系统碳总贮量分别为184.15,198.42和179.96 t·hm-2。生态系统碳贮存的空间分布排序为土壤层>植被层>凋落物层。植被层的碳贮量平均为44.76 t·hm-2,占平均总贮量的23.87%;凋落物层平均为3.48 t·hm-2,仅占1.86%,死地被物的碳贮量虽最小,但它是土壤-植物系统碳循环的联结库,对森林生态系统的碳循环起到重要的作用;林地土壤层(0~60 cm)碳贮量是相当可观的,平均为139.27 t·hm-2,占74.27%,可见林地土壤是碳的一个极重要的贮存库。
2.4 杉木年净固定量的初步估算森林生态系统生产力研究的主要内容之一是确定系统同化CO2的能力。根据各组分年净生产力及相应组分的碳素密度计算出有机碳年净固定量(方晰等,2002a)(表 8)。由表 8可看出,13年生杉木年净生产力为10.10 t·hm-2a-1,有机碳年净固定量为4.67 t·hm-2a-1,折合成CO2的量为17.13 t·hm-2a-1。森林不仅是陆地生态系统中最为重要的生物基因库,而且也是大气CO2的一个重要的汇。森林的破坏等于给大气增加一个重要的CO2供给源。因为森林砍伐,除了利用一部分有用的木材外,其他部分如树叶、枝、皮、根等可能被分解腐烂,或被当作燃料烧掉,通过这些途径向大气中释放CO2。保护好现有森林,每年可以吸收大气中一定量的CO2。由此可见,森林与全球大气CO2浓度的升降具有密切的关系,成为影响全球气候变化的一个重要因子,说明保护、恢复和发展森林植被具有非常重要的生态学意义。确定系统同化CO2的能力是森林生态系统生产力研究的主要内容之一。
|
|

13年生杉木人工林乔木层不同器官碳素各异,排列顺序为:叶>皮>干>根>枝,这与湖南会同的11年及14年生杉木(田大伦等,2004)基本一致。可见,地域及林龄差异并没影响各器官碳素含量的大小排序。各器官碳素含量平均值为497 g·kg-1,高于湖南会同的14年生杉木各器官碳素含量平均值(480.7 g·kg-1)(田大伦等,2004),低于湖南会同的23年生杉木各器官碳素含量平均值(463.2 g·kg-1)(陈楚莹等,2000),说明杉木生长区域及发育阶段的差异影响到乔木层各组分碳素含量的大小。随着林分发育,乔木层各组分碳素含量平均值呈现增加的趋势。广西14年生马尾松(Pinus massoniana)(方晰等,2003a)及16年生湿地松(Pinus elliottii)(方晰等,2003b)均表现出了不同的碳素含量大小规律,说明各器官碳素含量大小与树种密切相关。林下植被各组分碳含量明显低于乔木层相对应的各组分含量。灌木层碳含量大于草本层。植被层碳含量除了与植被种类有关外,还与环境条件密切相关。乔木层叶片处于林冠上部,有较长的日照时间和充足的光能,从而可以截获较多太阳能,有利于植物的光合作用,合成、积累较多的有机物,因而碳含量较高;由于凋落物中的部分有机物被分解,处于半分解状态的凋落物碳素含量明显低于未分解凋落物碳素含量。杉木林凋落物碳素平均含量高于樟树(Cinnamomum camphora)林下凋落物(403 g·kg-1)(雷丕锋等,2004),主要因为杉木林下凋落物有机碳的分解速率较阔叶林下的慢,碳累积较多;土壤各层次碳含量随着土层加深而降低。同一土层在各样区碳含量差异不明显。土壤碳素含量明显低于植被层及凋落物层。
杉木生态系统中碳库主要分为3个部分:植被层、凋落物层和土壤层。生态系统碳贮量平均为187.51 t·hm-2,低于广西中部14年生马尾松纯林生态系统碳贮量(235.0 t·hm-2)(方晰等,2003a)。乔木层碳贮量(43.0 t·hm-2)大于湖南会同11年生杉木乔木层碳贮量(34.53 t·hm-2)(方晰等,2002b)。杉木植被碳贮量为44.76 t·hm-2,小于我国森林植被平均碳贮量(57.07 t·hm-2)、暖性针叶林(47.97 t·hm-2)(周玉荣等,2000)及热带、亚热带针叶林63.7 t·hm -2(王绍强等,1999),高于温性针叶林(43.26 t·hm-2)(周玉荣等,2000)。植被层碳贮量主要取决于乔木层的生物量。杉木各器官中,3个样区树干平均碳贮量占乔木层平均总量的58.40%,若杉木林被采伐后,其树干可以被用作家具或其他林产品,其碳贮量可以保存下来,成为缓冲性质的林产品碳库,对调节地球环境中碳周转速率和周转量有重要意义(秦建华等,1997);其次,树根也占较大比例,占平均总贮量的20.09%,采伐后树根在土壤里保存可减少碳的流失;乔木枝、皮、叶碳平均总贮量占乔木层平均总贮量的21.51%。可见非木材的枝、叶、皮、根占较大比例,因此,采伐剩余物的处理和森林恢复更新对碳贮量的影响很大。如采伐后大量枝、叶、皮等作薪材或进行“炼山”,将导致大量有机质的分解,土壤表层有机质也迅速氧化分解,将导致大量的CO2排放(方晰等,2005;雷丕锋等,2004)。
凋落物层碳贮量较低,为3.48 t·hm-2,这主要是由于凋落物碳贮量很大程度取决于水热条件、地域特点等。一般纬度越高,分解条件越差,积累越多;反之,分解越迅速,积累较少(周玉荣等,2000)。
我国森林土壤平均碳贮量为193.55 t·hm-2,约是植被贮量的3.4倍(周玉荣等,2000)。广西大青山杉木林地土壤有机碳贮量(139.27 t·hm-2)及其与植被碳贮量的比值(3.1:1)均低于全国平均水平,这主要由于:一方面亚热带较好的水热条件利于植被生物量的累积;另一方面,亚热带土壤呼吸速率较大,加之植被对土壤中养分的大量吸收,造成土壤碳贮量相对较低(周玉荣等,2000)。土壤表层(0~20 cm)碳贮量贡献较大,占整个土壤有机碳贮量(0~60 cm)的40.76%,可见防止地表的水土流失,可有效保持土壤对碳的吸存。林下植被在维持地力、保持水土等方面有着重要的作用(方海波等,1998;Liu et al., 1995),以往林分抚育对林下植被破坏严重,不仅造成植被碳贮量的减少,而且加剧了地表径流对表土层的冲蚀,进而导致土壤碳散失。因此,在该区域保护林下植被对维持整个生态系统的碳汇有着非常重要的作用。
1999—2003年期间我国杉木林生态系统碳素现存量为2.866 Gt,1个龄级期后贮碳量将达到3.772 Gt,以平均每年90.63 Mt的积累速率递增,是一个贮碳潜力大,增长速率快的碳库(王兵等,2008)。初步估算13年生杉木人工林年净生产力为10.1 t·hm-2a-1,有机碳年净固定量为4.67 t·hm-2a-1,折合成CO2的量为17.13 t·hm-2a-1。湖南会同11年生杉木年净生产力为7.35 t·hm-2a-1,有机碳年净固定量为3.49 t·hm-2a-1(方晰等,2002b)。苏南地区27年生杉木生态系统1 hm2每年仅能吸收2.36 t碳(阮宏华等,1997)。热带山地雨林1 hm2每年能吸收3.818 t碳(李意德等,1998)。18年生樟树林1 hm2每年可吸收4.98 t碳(雷丕锋等,2004)。可见,杉木生态系统年固碳量较高。但由于受该区域极端温度的限制,杉木在中龄林前生长较快,中龄林后很快进入衰退阶段(王献溥等,2004),只能培育小径材。频繁的多代连栽导致杉木人工林生物量和土壤有机碳贮量减少(罗云建等,2006)。加之,极限区域对全球变化的敏感性,使得该区域杉木人工林栽培面积及净初级生产力(NPP)也呈现减少的趋势(刘世荣等,2005)。因此,该区域杉木人工林固碳潜力会下降。研究杉木极限分布区生态系统固碳量,对评价及管理杉木生态系统碳汇功能具有较为深远的意义。该生态系统碳动态及平衡有待于进一步研究。
陈楚莹, 廖利平, 汪思龙. 2000. 杉木人工林生态系统碳素分配与贮量的研究. 应用生态学报, 11(增1): 175-178. |
方海波, 田大伦, 康文星. 1998. 杉木人工林间伐后林下植被生物量的研究. 中南林学院学报, 18(1): 6-9. |
方精云, 陈安平. 2001. 中国森林植被碳库的动态变化及其意义. 植物学报, 43(9): 967-973. DOI:10.3321/j.issn:1672-9072.2001.09.014 |
方晰, 田大伦, 项文化. 2002a. 速生阶段杉木人工林碳素密度、贮量和分布. 林业科学, 38(3): 14-19. |
方晰, 田大伦, 项文化. 2002b. 第二代杉木中幼林生态系统碳动态与平衡. 中南林学院学报, 22(1): 1-6. |
方晰, 田大伦, 胥灿辉. 2003a. 马尾松人工林生产与碳素动态. 中南林学院学报, 23(2): 11-15. |
方晰, 田大伦, 项文化, 等. 2003b. 不同密度湿地松人工林中碳的积累与分配. 浙江林学院学报, 20(4): 374-379. |
方晰, 田大伦, 项文化, 等. 2005. 杉木人工林林地土壤CO2释放量及其影响因子的研究. 林业科学, 41(2): 1-6. |
康冰, 刘世荣, 张广军, 等. 2006. 广西大青山南亚热带马尾松、杉木混交林生态系统碳素积累和分配特征. 生态学报, 26(5): 1320-1328. DOI:10.3321/j.issn:1000-0933.2006.05.004 |
李意德, 吴仲民, 曾庆波, 等. 1998. 尖峰岭热带山地雨林生态系统碳平衡的初步研究. 生态学报, 18(4): 371-378. |
雷丕锋, 项文化, 田大伦, 等. 2004. 樟树人工林生态系统碳素贮量与分布研究. 生态学杂志, 23(4): 25-33. DOI:10.3321/j.issn:1000-4890.2004.04.006 |
刘世荣, 温远光. 2005. 杉木生产力生态学. 北京: 气象出版社.
|
罗云建, 张小全. 2006. 多代连栽人工林碳贮量的变化. 林业科学研究, 19(6): 791-798. DOI:10.3321/j.issn:1001-1498.2006.06.020 |
裴志永, 欧阳华, 周才平. 2003. 青藏高原高寒草原碳排放及其碳循环迁移过程研究. 生态学报, 23(2): 231-236. DOI:10.3321/j.issn:1000-0933.2003.02.002 |
秦建华, 姜志林. 1997. 森林在大气碳平衡中的作用. 世界林业研究, (4): 18-25. |
阮宏华, 姜志林, 高苏铭. 1997. 苏南丘陵主要森林类型碳循环研究:含量与分布规律. 生态学杂志, 16(6): 17-21. DOI:10.3321/j.issn:1000-4890.1997.06.004 |
田大伦, 方晰. 2004. 湖南会同杉木人工林生态系统的碳素含量. 中南林学院学报, 24(2): 1-5. DOI:10.3969/j.issn.1673-923X.2004.02.001 |
王绍强, 周成虎, 罗承文, 等. 1999. 中国陆地自然植被碳量空间分布探讨. 地理科学进展, 18(3): 238-244. DOI:10.3969/j.issn.1007-6301.1999.03.007 |
王献溥, 郭柯, 田新智. 2004. 广西杉木林的分类、分布和演替. 植物资源与环境学报, 13(1): 43-47. DOI:10.3969/j.issn.1674-7895.2004.01.010 |
王兵, 魏文俊, 李少宁, 等. 2008. 中国杉木林生态系统碳储量研究. 中山大学学报:自然科学版, 47(2): 93-98. |
中国农业化学专业委员会. 1983. 土壤农业化学常规分析方法. 北京: 科学出版社, 272-273.
|
周玉荣, 于振良, 赵士洞. 2000. 我国主要森林生态系统碳贮量和碳平衡. 植物生态学报, 24(5): 518-522. DOI:10.3321/j.issn:1005-264X.2000.05.002 |
Artuso F, Chamard P, Piacentino S, et al. 2009. Influence of transport and trends in atmospheric CO2 at Lampedusa. Atmospheric Environment, 43(19): 3044-3055. DOI:10.1016/j.atmosenv.2009.03.027 |
Christopher S G, Robert B J. 2009. Risks to forest carbon offset projects in a changing climate. Forest Ecology and Management, 257: 2209-2216. DOI:10.1016/j.foreco.2009.03.017 |
IPCC. 2001. Climate Change 2001: the science of climate change. Summary for policy-markers, a report of working group Ⅰ of the intergovernmental panel on climate change. Geneva, Switzerland, 1-98.
|
Liu S R. 1995. Nitrogen cycling and dynamic analysis of man-made larch forest ecosystem. J Plant Soil, 168/169: 391-397. DOI:10.1007/BF00029352 |
Norby R J, Luo Y. 2006. Evaluating ecosystem responses to rising atmospheric CO2 and global warming in a multi-factor world. New Phytologist, 162(2): 281-293. |
Onigkeit J, Sonntag M, Alcamo J. 2000. Carbon plantations in the IMAGE model-model description and scenarios. Center for Environmental Systems Research, University of Kassel, Germany, 35-113.
|
Tans P P. 2006. How can global warming be traced to CO2. Scientific American, 295(6): 124. DOI:10.1038/scientificamerican1206-124 |