 2009, Vol. 45
2009, Vol. 45文章信息
- 周兰英, 王永清, 张丽, 胡泽明.
- Zhou Lanying, Wang Yongqing, Zhang Li, Hu Zeming
- 46种杜鹃花属植物表型性状的数量分类
- Mathematic Classification of 46 Species in Rhododendron with the Morphologic Characters
- 林业科学, 2009, 45(8): 67-75.
- Scientia Silvae Sinicae, 2009, 45(8): 67-75.
-
文章历史
- 收稿日期:2008-09-22
-
作者相关文章



2. 四川省会理县林业局 会理 615100
2. Huili County Forestry Bureau, Sichuan Province Huili 615100
杜鹃花属(Rhododendron)植物多具有极高的观赏价值, 是世界著名观赏花卉; 从中提取的活性成分可抗菌消炎、降低血压(钟国华等, 2000); 杜鹃根系发达, 枝条密集, 对维系山区生态环境具有重要作用.中国西南是世界最大的杜鹃分布中心和多度中心, 野生杜鹃资源丰富, 具有很高的开发利用价值, 但由于杜鹃花属植物种类多、分布广、变异大, 分类问题十分复杂.传统的形态分类往往进行定性描述, 缺乏定量分析, 带有一定的主观色彩, 在系统划分和某些种类的归属上长期存在分歧, 影响了杜鹃花属植物的开发利用.
传统的形态研究与计算机技术结合, 形成了数量分类学.数量分类使生物分类学正逐渐从定性描述向定量的、精确的高水平发展.近年来, 数量分类学已被应用于品种分类(赵冰等, 2007)、科属下种间分类(闫双喜等, 2002; 彭焱松等, 2007; 张汉尧等, 2005)、系统学研究(闫双喜等, 2004; Eduardo et al., 2002)等.建立在数量分类基础上的主成分分析是一种掌握主要矛盾的统计分析方法, 它可以从多元事物中揭示生命现象的主要方面, 现已应用于生态学(李景国, 2007; 巫锡柱等, 2007)、林业(邢世岩等, 2000; 李周岐等, 2000; 钟伟华等, 1999; 赵亮等, 2007)等领域的研究, 成为研究生物学及其边缘学科的重要工具.
有关杜鹃花属植物数量分类的研究迄今在国内外尚未见报道.本研究对四川西南山地46种野生杜鹃花属植物的表型性状进行系统聚类分析和主成分分析, 旨在确定重要分类指标, 揭示主要性状之间的内在联系及各性状所占有的信息比例, 为进一步研究种间亲缘关系及本属植物的开发利用提供科学依据.
1 材料与方法 1.1 材料来源本研究的46种杜鹃中31种从四川西部山地采集, 海拔2 020~3 435 m, 另15种从中国科学院华西亚高山植物园都江堰龙池杜鹃园采取.按Sleumer(1949; 1980)分类系统, 分属3个亚属, 详见表 1.
|
|
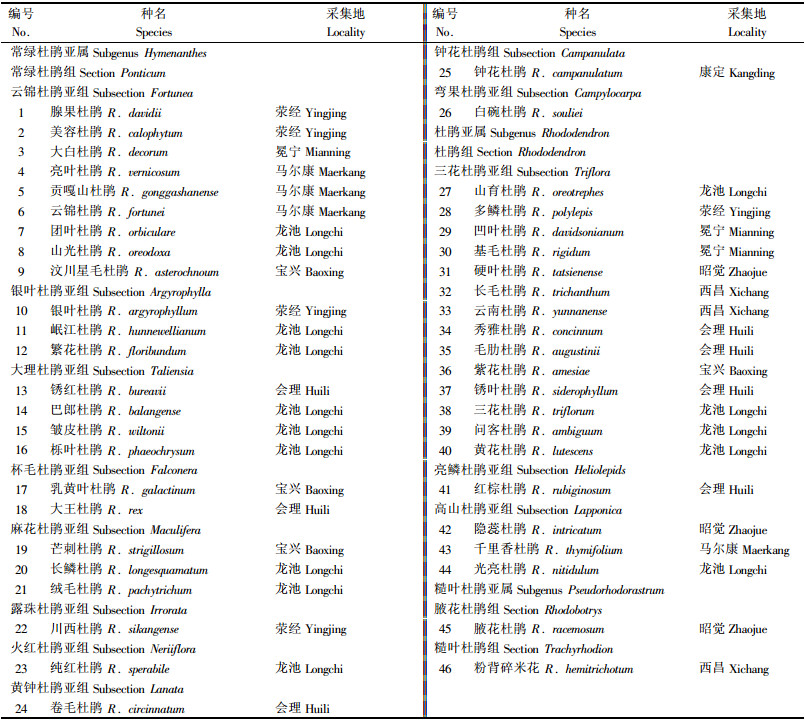
将采集到的46种杜鹃作为46个分类运算单位(OTU).通过对杜鹃的实地调查、标本的观察测量, 获取有关杜鹃种各性状的原始记录.经过对原始记录进行分析整理, 筛选出在杜鹃花分类中常用的44个表型性状进行编码, 其中二元性状18个, 编码0, 1;多元性状13个, 编码0, 1, 2…; 直接数量性状11个, 测定值或计数值即为编码; 转换数量性状2个, 分别是叶柄相对长和花梗相对长, 其中叶柄相对长为叶柄长与叶片长之比, 花梗相对长为花梗长与花冠长之比.一般而言, 叶片越长, 叶柄也越长, 花梗长和花冠长也基本呈正相关, 用相对长度可避免计算过程中叶长和冠长的二次影响.各性状编码详见表 2.
|
|
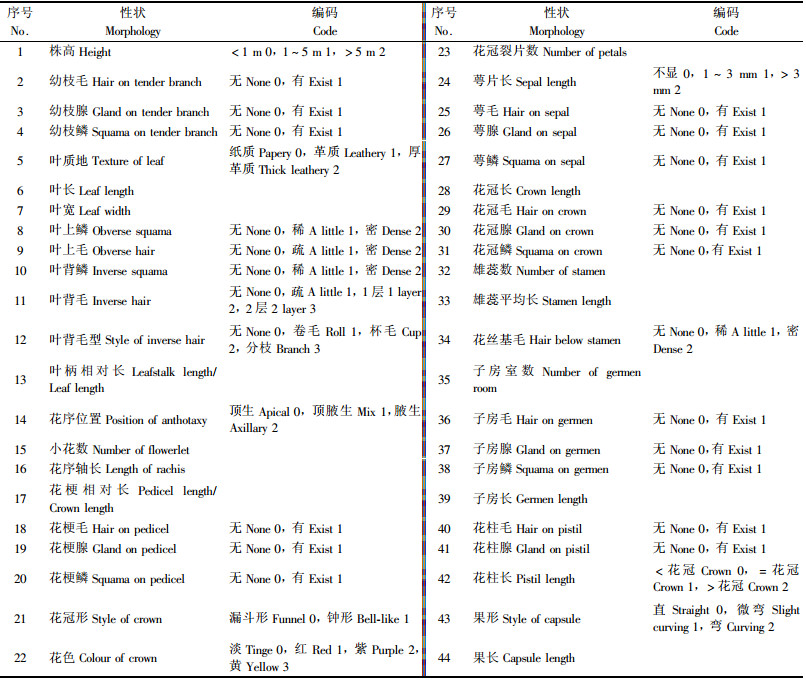
叶长、宽、叶柄长从每份标本上随机抽取5~20片叶测定后计算平均值; 花冠长宽、花梗长、子房长、雄蕊长每份标本上随机抽5朵小花测定后计算平均值; 果长每份标本上随机抽5个果测定后计算平均值, 长度单位均为mm; 雄蕊数和子房数为多次计数结果.每种植物测定标本数3个以上.
1.2.3 数据处理及计算方法数据在进行运算之前, 以标准差标准化法(徐克学, 1999)对原始数据进行处理, 以标准化数据计算各OTU之间的距离系数.
1.2.4 聚类方法用SPSS 10.0 for windows采用平均欧氏距离平方系数作Q聚类, 分析杜鹃种间的表型性状相似程度; 用相同软件采用相关系数作R聚类, 分析性状间的相关程度. 2种聚类均采用类间平均连锁法.根据聚合过程中ΔDK和ΔRK值的跳变情况分别确定Q聚类和R聚类的结合线.
1.2.5 主成分分析计算各性状间相关系数得相关系数矩阵; 根据成分提取结果确定主成分; 以李朝锋等(2007)的方法计算特征向量和主成分向量; 以主成分1向量为横坐标, 主成分2向量为纵坐标作散点图.
2 结果与分析 2.1 聚类结果分析 2.1.1 Q聚类46种杜鹃按欧氏平方距离系数由小到大进行聚合, 聚合过程及距离见表 3.从表 3可见, 最大的跳变出现在聚合的第43步, 其次是第35步和第21步, 据此确定3条结合线的位置.
|
|
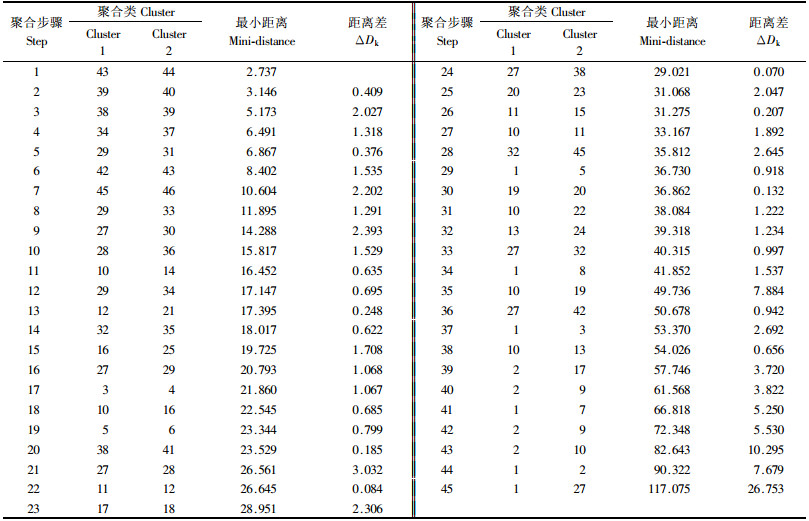
从聚类图(图 1)上可见, 结合线L1将46种杜鹃划分为2个大类群, 第1类群(Ⅰ)共20种, 包括了全部有鳞类杜鹃; 第2类群(Ⅱ), 由常绿杜鹃亚属的26个种组成.

|
图 1 46种杜鹃聚类图 Figure 1 Dendrogram of 46 Rhododendron |
结合线L2将Ⅰ类群的20种有鳞杜鹃全部划为1类; Ⅱ类群被分为A, B, C, D 4部分: A部分是云锦杜鹃亚组的7个种, B部分只有1种白碗杜鹃; C部分包括14个种; D部分由杯毛杜鹃亚组的2个种和云锦杜鹃亚组中的美容杜鹃和汶川星毛杜鹃组成, 均为乔木, 叶极大, 小花数多.
结合线L3将有鳞的Ⅰ类群划分为a, b, c 3个小组: a小组只有高山杜鹃亚组的3个种, 其植株、叶片、花冠等均非常小; b小组由三花杜鹃亚组的12个种和亮鳞杜鹃亚组的红棕杜鹃组成, 均表现幼枝、叶背无毛, 花冠较大, 子房较长; c小组的4种杜鹃共同点是幼枝、叶两面、花萼、子房等明显有毛. L3将ⅡA类群进一步划分为a, b, c 3个小组, 反映的是云锦杜鹃亚组内的分化; Ⅱ B类群只有1个种; Ⅱ C类群被L3划分为a, b, c 3个小组, 其中c小组有9个种, 分属银叶杜鹃亚组、大理杜鹃亚组、钟花杜鹃亚组、露珠杜鹃亚组; Ⅱ D类群被分为2个小组.每小组种类都较少, 说明常绿杜鹃亚属内种间表型变异幅度较大.
通过对欧氏平方距离系数矩阵的整理和计算, 常绿杜鹃亚属内26个种间平均距离72.310, 有鳞类20种杜鹃种间平均距离35.264, 表明有鳞类杜鹃在表型上变异相对较小; 46种杜鹃平均欧氏平方距离为90.00, 最大达229.938, 距离系数差异巨大, 意味着杜鹃花属内种间表型性状分化十分剧烈.
2.1.2 R聚类1)性状分类 各性状按相关系数由大到小逐步聚合, 聚合结果见性状聚类树系(图 2).由图 2可见, 结合线将44个分类性状划分为4类. A类共12个性状, 其中7个性状与鳞片有关, 且各部位有鳞与否相关紧密, 其余5个性状有4个表现花器特征; B类只有花冠有毛与否1个性状; C类有6个性状, 全部与毛有关; D类性状最多, 达25个, 又可再划分为2个小类, 第1小类主要与腺体有关, 第2小类基本全是数量性状.

|
图 2 杜鹃性状聚类图 Figure 2 Dendrogram of morphology |
2)性状间的相关性 从R聚类所得性状相关系数矩阵可见, 部分性状之间相关程度较高, 其中花萼、子房、幼枝、花梗有无鳞片4性状间相关系数皆为1, 说明这些性状相伴发生; 叶背鳞片有无与该4性状相关系数为0.9330, 也几乎同时出现; 鳞片的出现与株高、叶、花、子房、果等的长度呈明显负相关, 绝对值均在0.8以上, 反映了有鳞类杜鹃普遍较常绿类杜鹃矮小且各器官也明显较小; 子房室数、小花数与花冠、子房、总梗长等性状的相关系数在0.75以上, 反映了花器构造有一定内部联系; 此外, 凡是反映器官尺寸的数量性状相关系数基本都在0.7以上, 表明各器官外部尺寸基本同步变化; 花冠有无毛或鳞片或腺体、花丝基部有毛与否、花冠裂片数、花柱相对长、有无附属物、雄蕊长、花萼裂片长等性状与其他性状相关系数几乎全在±0.4以下, 说明这些性状的出现基本没有规律可言, 在种以上分类阶元的划分中作用不大.
2.2 主成分分析R聚类结果已表明部分性状间相关系数很高, 存在信息重叠; 部分性状的出现基本没有规律可言, 在种以上类群的划分中没有明显作用, 因此有必要作主成分分析.通过主成分分析, 可以排除重叠和分类意义不大的性状, 使重要性状突显出来.
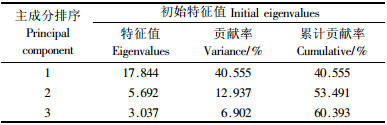
2.2.1 主成分提取成分提取分析(表 4)是对44个分析特征(观测性状)综合计算后, 按其在分类中的作用和类别重新划出的特征成分各自的特征值和贡献率.从表 4可见, 第1主成分所占达信息量就达40.555%, 前3个成分占信息总量的60.393%, 而其余41个成分所提供的信息量仅39.607%, 说明在杜鹃分类研究中存在重要性状, 有必要作主成分分析.
|
|
将初始因子载荷矩阵和性状抽提系数表综合起来, 得到用于杜鹃分类的44个特征性状对应的主成分载荷量、总载荷量和性状重要性排序, 详见表 5.
|
|
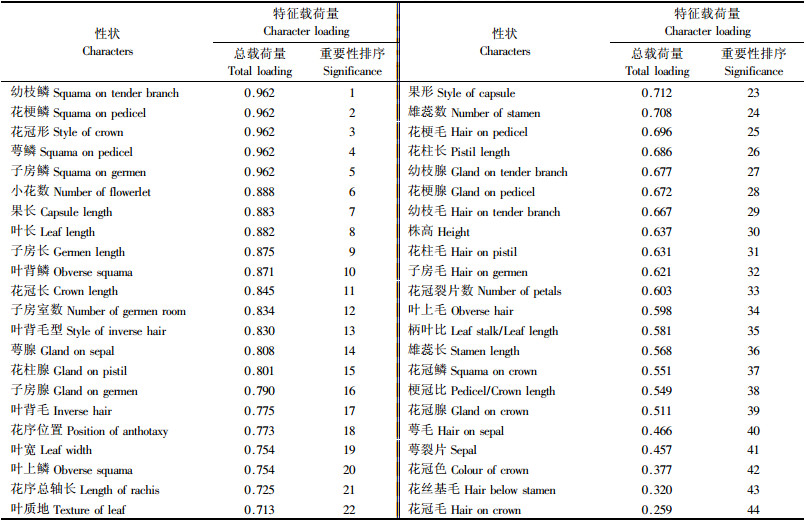
从表 5可知, 第1主成分在幼枝鳞、叶背鳞、花梗鳞、萼鳞、子房鳞5个性状上均呈现很高的负向载荷量, 绝对值均在0.9以上, 在叶上是否有鳞上载荷量稍低, 为-0.807.与鳞片有无相反, 第1主成分与叶长、叶宽、花冠长、子房长、果长等性状均表现较高的正向载荷量, 全部超过0.75.这些性状的信息载荷很高, 表明第1主成分主要反映植株是否被鳞以及叶、花、果、子房等器官的外部尺寸, 也说明这些性状对杜鹃分类很重要.载荷量的分化同时也明显反映出第1主成分将46种杜鹃分为2大类群:较大型的无鳞类和较小型的有鳞类; 第2主成分在幼枝、花冠、花萼、叶片是否具有腺体或毛被等性状上载荷量较高, 说明这些性状对杜鹃分类较为重要.
从性状排序(表 5)上可见, 对杜鹃分类最重要的外表是否具鳞或腺体或毛被, 其次是叶、花、子房、果的大小, 其总载荷量均在0.8以上, 重要性占据了所有性状的前15位; 重要性排后10位的分别是叶柄相对长、雄蕊长、花冠鳞、花梗相对长、花冠腺、花萼毛、萼裂片长、花冠色、花丝基毛、花冠毛, 其中萼片是否具毛、萼裂片长短、花冠颜色、花丝基部毛、花冠毛在前3个主成分的总载荷量中重要性最不明显, 总载荷量都在0.5以下, 对分类贡献最小.
2.2.3 主成分向量及作用以第1, 2主成分向量分别为横坐标和纵坐标作散点图(图3).从图3上可见, 46种杜鹃被明显分为2个大类, 左边全是有鳞类杜鹃, 右边则全是无鳞的常绿类杜鹃.在有鳞杜鹃类群中, 最左边的是糙叶杜鹃亚属的2个种, 虽与杜鹃亚属比较接近, 但并未混淆.仅用2个主成分就能比较有效地对亚属进行分类, 说明主成分提取方法可行, 结果可信.

Sleumer(1949)将花序腋生的有鳞类杜鹃单独划为1个糙叶杜鹃亚属, 但多数学者主张将其划入杜鹃亚属杜鹃组中作亚组处理(Chamberlain et al., 1996; Cullen, 1980; Philipson et al., 1982).长期以来, 糙叶类杜鹃的归属一直是争论的焦点之一.从表型性状上看, 糙叶杜鹃亚属与杜鹃亚属之间确无太大差异, 在本研究中, 2亚属间的平均欧氏距离为47.16, 而同属杜鹃亚属杜鹃组的高山杜鹃亚组与三花亚组之间平均距离为48.76, 两者比较, 看不出亚属间的差异.但一些研究结果表明, 在某些方面2亚属间又存在明显差异. 高连明等(2000)采用水平淀粉凝胶电泳分析, 显示糙叶杜鹃亚属内的3种杜鹃遗传相似性最高, 与其他杜鹃的遗传相似性较低; 熊子仙(1996)通过对糙叶杜鹃亚属的6个种叶解剖特征观察, 提出该亚属植物具有角质层发达、外层细胞小、内层细胞大、叶主脉为周韧维管束等特征, 认可Sleumer(1949)将其划分为1个亚属的分类系统; 花粉电镜扫描结果也显示2亚属有一定差异(周兰英等, 2008).因此, 糙叶类杜鹃分类地位的合理确定, 尚需要对更多种类和在不同领域作更深入的研究.
3.2 亚组及种间关系本研究中, 亮鳞杜鹃亚组的红棕杜鹃与三花杜鹃亚组的平均距离为28.958, 而三花杜鹃亚组内种间平均距离为26.640, 二者并未表现明显差异.由于这2个亚组形态差异本身很小, 亚组划分界线不清晰, 因此可以考虑合并为1个亚组.
在传统的形态分类中, 美容杜鹃和汶川星毛杜鹃被划分在常绿杜鹃亚属下云锦杜鹃亚组中, 但在本研究Q聚类中, 该亚组的另7种杜鹃在较小距离内聚合在一起, 美容杜鹃和汶川星毛杜鹃却与之相距较远, 最终与杯毛杜鹃亚组的大王杜鹃和乳黄叶杜鹃聚合为1类.按分类检索表, 云锦杜鹃亚组的主要特征是幼枝无毛, 稀有腺体或绒毛; 叶较小, 长不到20 cm; 子房有腺体(何明友等, 1994).而这2种杜鹃与同组的其他7种杜鹃有明显区别:幼枝有毛; 叶平均长超过23 cm, 大的超过30 cm; 子房和花柱无腺体, 所以它们是否应归入云锦杜鹃亚组值得商榷.
3.3 分类性状的选择R聚类和主成分分析结果都揭示分类性状并非越多越好, 一部分性状如雄蕊长、花冠有无附属物、花梗长、花萼毛、花冠颜色、花丝基部有毛与否、花冠裂片数、花柱长、有无附属物等性状在属下分类等级上作用很小, 在杜鹃分类时应选择重要性状, 更多地注意有无鳞片、有无毛被及类型、叶、花、子房、果的大小等, 避免过多次要性状对分类的干扰.
高连明, 张长芹, 王中仁. 2000. 9种杜鹃花属植物的遗传分化研究. 广西植物, 20(4): 377-382. DOI:10.3969/j.issn.1000-3142.2000.04.019 |
何明友, 方明渊, 胡文光, 等. 1994.中国植物志: 第57卷2分册.北京: 科学出版社, 2-3.
|
李朝锋, 杨中宝. 2007. SPSS主成分分析中的特征向量计算问题. 统计教育, (3): 10-11. |
李景国. 2007. 重庆三峡库区生态经济区农业和农村经济状况评价. 安徽农业科学, 35(26): 8390-8392. DOI:10.3969/j.issn.0517-6611.2007.26.149 |
李周岐, 王章荣. 2000. 鹅掌揪属种间杂种F1与亲本花果数量性状的遗传变异分析. 林业科学研究, 13(3): 290-294. DOI:10.3321/j.issn:1001-1498.2000.03.011 |
彭焱松, 陈丽, 李建强, 等. 2007. 中国栎属植物的数量分类研究. 武汉植物学研究, 25(2): 149-157. DOI:10.3969/j.issn.2095-0837.2007.02.008 |
巫锡柱, 晏路明. 2007. 脆弱生态环境的综合评判物元模型研究. 中国生态农业学报, 15(3): 138-141. |
邢世岩, 倪国祥, 张运吉, 等. 2000. 银杏叶数量性状的遗传分析. 林业科学, 36(5): 47-53. DOI:10.3321/j.issn:1001-7488.2000.05.008 |
熊子仙. 1996. 糙叶杜鹃亚属叶解剖特征与组的划分. 云南师范大学学报, 16(4): 62-63. |
徐克学. 1999. 生物数学. 北京: 科学出版社, 51-70.
|
闫双喜, 赵勇. 2002. 中国黄杨属植物数量分类的研究. 生物数学学报, 17(3): 380-383. DOI:10.3969/j.issn.1001-9626.2002.03.021 |
闫双喜, 杨秋生, 王鹏飞, 等. 2004. 中国部分地区种子植物区系亲缘关系的研究. 武汉植物学研究, 22(3): 226-230. DOI:10.3969/j.issn.2095-0837.2004.03.008 |
张汉尧, 刘小珍, 孙茂胜, 等. 2005. 竹子形态标记聚类分析研究. 竹子研究汇刊, 24(1): 25-28. DOI:10.3969/j.issn.1000-6567.2005.01.005 |
赵冰, 雒新艳, 张启翔. 2007. 蜡梅品种的数量分类研究. 园艺学报, 34(4): 947-954. DOI:10.3321/j.issn:0513-353x.2007.04.023 |
赵亮, 陶红军. 2007. 湖北省林业综合生产能力研究. 林业经济问题, 27(3): 272-275. DOI:10.3969/j.issn.1005-9709.2007.03.018 |
钟国华, 胡美英. 2000. 杜鹃花科植物活性成分及作用机制研究进展. 武汉植物学研究, 18(6): 509-514. DOI:10.3969/j.issn.2095-0837.2000.06.012 |
钟伟华, 何昭珩. 1999. 主成分聚类在火炬松种子园疏伐中的运用. 福建林学院学报, 19(1): 33-36. DOI:10.3969/j.issn.1001-389X.1999.01.010 |
周兰英, 王永清, 张丽. 2008. 26种杜鹃花属植物花粉形态及分类学研究. 林业科学, 44(2): 55-63. DOI:10.3321/j.issn:1001-7488.2008.02.009 |
Chamberlain D F, Hyam R, Argent G, et al. 1996. The genus Rhododendron, its classification and synonymy. Edinburgh: Royal Botanic Garden Edinburgh.
|
Cullen J. 1980. A revision of Rhododendron L. Subgenus Rhododendron Section Rhododendron and Pogonanthum. Notes Royal Botanic Garden Edinburgh, 39(1): 1-208. |
Eduardo L, Borb A, Gerorge J, et al. 2002. Floral and vegetative morphometrics of five Pleurothallis (Orchidaceae) species: Correlation with taxonomy, phylogeny, genetic variability and pollination systems. Annals of Botany, 90: 219-230. DOI:10.1093/aob/mcf168 |
Philipson M N, Philipson W R. 1982. A preliminary synopsis of the genus Rhododendron. Notes Royal Botanic Garden Edinburgh, 40(1): 225-227. |
Sleumer H. 1949. Ein system dergattung Rhododendron L.. Bot Jahrb Syst, 74: 511-553. |
Sleumer H.1980. Past and present taxonomic systems of Rhododendron based on macromorphological characters//Luteyn J L, O'Brien M E. Contributions toward a classification of Rhododendron. New York: New York Botanical Garden, 19-26.
|