 2009, Vol. 45
2009, Vol. 45文章信息
- 臧德奎, 陈红, 郑林, 郭先锋.
- Zang Dekui, Chen Hong, Zheng Lin, Guo Xianfeng
- 木瓜属优良品种亲缘关系的AFLP分析
- Analysis on the Phylogenetic Relationship of Elite Cultivars in Chaenomeles Using AFLP
- 林业科学, 2009, 45(8): 39-43.
- Scientia Silvae Sinicae, 2009, 45(8): 39-43.
-
文章历史
- 收稿日期:2008-06-02
-
作者相关文章


木瓜属(Chaenomeles)是重要的观赏和药用植物, 有5种和1个杂交种, 即贴梗海棠(C. speciosa)、木瓜海棠(C. cathayensis)、木瓜(C. sinensis)、西藏木瓜(C. tibetica)、日本木瓜(C. japonica)和傲大贴梗海棠(C. × superba)(俞德浚, 1974; Brickell, 1996). 我国是木瓜属植物的分布中心和栽培中心, 品种资源丰富, 对其进行遗传多样性及亲缘关系的研究, 有助于了解品种起源和演化, 为杂交育种和品种选育及核心种质构建提供依据. 前人对木瓜属植物的研究集中在品种调查和形态学标记等方面, 赵红霞(2003)、王嘉祥(2004)、张茜等(2005)先后探讨过木瓜属栽培品种的形态分类, 但品种的植物学归属和品种划分存在较大争议. AFLP分子标记具有带型丰富、用样量少、灵敏度高、快速高效等特点, 已成为遗传多样性和品种分类研究的有效手段(Bohn et al., 1999; Dong et al., 2000; Tomkins et al., 2001; 杨朝东等, 2005; 沈镝等, 2005; 黄久香等, 2008). 本研究利用荧光AFLP技术, 对木瓜属主要栽培品种进行评价, 从分子水平上探讨品种间的遗传多样性及亲缘关系, 以期为核心种质构建和育种亲本选配提供理论指导.
1 材料与方法 1.1 材料与来源供试木瓜属品种共29份, 为本属常见的优良品种, 分别来自山东农业大学木瓜种质资源圃(泰安)和山东临沂市河东区汤河镇. 29份种质材料的编号、名称及来源见表 1.
|
|

采用改进的CTAB法(Fang et al., 1992)从硅胶干燥的幼叶中提取总DNA.
1.2.2 AFLP分析利用北京鼎国生物技术公司的试剂盒及其操作指南进行. 各样品基因组DNA的酶切和接头的连接在同一反应中进行. 在20 μL反应体系中含有DNA模板4 μL (50 ng·μL-1), Adapter 1 μL, EcoRⅠ/MseⅠ (4 U·μL-1) 2 μL, 10 × Reaction buffer 2.5 μL, ATP (10 mmol·L-1) 2.5 μL, 4 Ligase (4 U·μL-1) 1 μL, AFLP-Water 7 μL. 将上述混合液在0.5 mL离心管中混匀离心数秒, 37 ℃保温5 h, 8 ℃保温4 h, 4 ℃过夜.
用预扩增引物组合进行扩增. 反应体系为25 μL. Pre-ampmix 1 μL, dNTPs 1 μL, 10×PCR buffer 2.5 μL, Taq DNA polymerase (2 U·μL-1) 0.5 μL, AFLP-Water 18 μL. 预扩增反应程序为: 94 ℃变性2 min; 94 ℃变性30 s, 56 ℃复性30 s, 72 ℃延伸80 s, 循环30轮; 72 ℃延伸5 min. 预扩增产物稀释20倍, -20 ℃保存备用.
用荧光标记的经过筛选的8对引物(表 2)进行选择性扩增. 反应体系为25 μL. 含预扩增稀释样品2 μL, dNTPs 2.5 μL, 10×PCR buffer 0.5 μL, EcoRⅠ引物1 μL, MseⅠ引物(荧光标记) 1 μL, Taq DNA polymease (2 U·μL-1) 0.5 μL, AFLP-Water 17.5 μL.
|
|
扩增反应按下列参数PCR循环: 94 ℃ 2 min; 一轮扩增参数: 94 ℃ 30 s, 65 ℃ 30 s, 72 ℃ 80 s; 以后每轮循环退火温度递减0.7 ℃或1 ℃, 扩增12轮; 接着按下列参数扩增23轮: 94 ℃ 30 s, 55 ℃ 30 s, 72 ℃ 80 s; 最后延伸72 ℃ 5 min.
1.2.3 电泳与数据处理取选择性扩增后的样品在ABI 377自动测序仪上电泳分离检测, 得到AFLP的DNA指纹图谱. 利用GeneScan 3.1软件将8对荧光引物产生的电泳胶图转换为(0, 1)矩阵. 根据PCR扩增的结果, 有带计为1, 无带则计为0, 依据CGS=m/(m+n)计算遗传相似系数, 式中m为基因型间共有带的数目, n为差异带的数目. 采用NTSYS-PC2.11版软件中的DICE法计算样本间的相似系数, 按非加权类平均法(UPGMA)进行聚类.
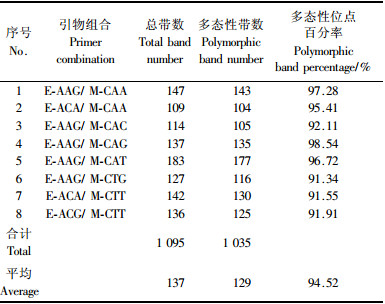
2 结果与分析 2.1 扩增产物的多态性利用从64对引物中筛选出的8对AFLP引物对供试品种的基因组DNA进行片段长度多态性扩增, 获得了较好的扩增结果(表 2). 共扩增出1 095条谱带(70~500 bp), 其中1 035条具有多态性, 占94.52%, 平均每对引物扩增出137条可统计的带, 其中129条具有多态性. 带数最多的是引物组合E-AAG/ M-CAT, 为183条, 其中多态性条带177条. 可见, AFLP检测木瓜属品种遗传多样性的效率很高, 也充分体现了该类植物丰富的遗传多样性.
从DNA分子水平上而言, 遗传多样性越高, 表明其遗传背景越复杂(Aggarwal et al., 1999), 高达94.52%的多态性表明木瓜属植物的遗传背景非常复杂, 我国木瓜属栽培品种形成较早, 在遗传进化过程中基因组DNA发生了丰富的变异.
2.2 木瓜属品种的亲缘关系29个木瓜属品种的相似系数在0.465 0~0.833 9之间(表 3). 其中, ‘凤凰木’与‘夕照’的相似系数最大, 为0.833 9, 表明2个品种间的亲缘关系最近; ‘红星’与‘长俊’的相似系数最小, 为0.465 0, 表明二者的亲缘关系最远. AFLP能将所有的材料分辨开, 品种间区分率达到100%. 另外, 在供试品种中, ‘长俊’与其他品种的平均相似系数最小, 为0.539 0, 而‘报春’与其他品种间的平均相似系数最大, 为0.715 5, 这表明材料间的相似系数变化较大, 供试品种遗传变异大.
|
|

采用UPGMA进行聚类分析, 结果见图 1. 从聚类图可以看出, 以相似系数0.62为标准可以将29个品种分为4类. 第1类由5个品种组成, 包括‘红霞’、‘醉杨妃’、‘罗扶’、‘一品香’和‘长俊’, 均属于木瓜海棠, 植株高大, 除了‘醉杨妃’外均为单瓣花、子房发育正常、果实大. 该类品种与其他3类亲缘关系较为疏远, 最先聚为一类. 在相似系数为0.68时又可分为2组, 其中果实较大、呈细长纺锤状的‘长俊’单独聚为第1小组; 其余3个品种聚为第2组, 均果实较小、呈圆形或圆柱形. 这表明‘长俊’与该种的其他品种关系较为疏远, 而‘红霞’和‘一品香’亲缘关系最近. 而且, 结合形态特征的相似性, 可以推断‘醉杨妃’系由‘一品香’通过芽变而产生.

|
图 1 29个木瓜属优良品种的AFLP聚类图 Figure 1 Dendrogram of 29 elite cultivars in Chaenomeles based on AFLP |
第2类由7个品种组成, 即‘单白’、‘碧雪’、‘单粉’、‘日落’、‘矮红’、‘日本红’和‘四季红’, 除‘碧雪’外, 均为日本木瓜的品种. 在相似系数0.68时, 该类也可分为2组, 其中, ‘单白’、‘碧雪’、‘单粉’和‘日落’单独相聚, 这几个品种在形态上表现为植株矮小, 具有日本木瓜的典型特征, 而且花色较浅, 为乳白色或淡粉色, 花朵较小, 全开时杯形; ‘矮红’、‘四季红’和‘日本红’聚为一组, 这3个品种比较特别, 花均为红色、重瓣, 直径较大, 植株也较前一组稍高大, 代表着日本木瓜内的1个特殊类型.
第3类由11个品种组成, 包括‘长寿乐’、‘绿宝石’、‘紫衣’、‘大富贵’、‘红宝石’、‘复长寿’、‘沂红’、‘粉蝶’、‘红舞’、‘沂橙’和‘骄阳’. 该类品种均属于傲大贴梗海棠, 来源于日本木瓜和贴梗海棠的杂交, 植株大多较贴梗海棠矮小. 相对于其他种类, 本类品种聚合较早, 品种间的遗传多样性差别较小. 该类在相似系数0.73处分为3组, 第1组由‘红舞’、‘沂橙’、‘骄阳’、‘复长寿’组成, 花红色或浅橙色, 花瓣轮数较少, 并有显著的雄蕊瓣化现象; 第2组包括‘绿宝石’和‘粉蝶’, 花重瓣, 花色浅, 白色或粉红色; 第3组包括‘长寿乐’、‘紫衣’、‘大富贵’、‘红宝石’和‘沂红’, 均为重瓣花, 红色或鲜红色.
第4类由6个品种组成, 包括‘猩红与金黄’、‘报春’、‘多彩’、‘凤凰木’、‘红星’和‘夕照’, 除‘猩红与金黄’外, 均属于贴梗海棠. 在相似系数为0.68时该类又可分为2组, ‘多彩’因花复色, 花冠呈红、白、粉等色, 与其他品种迥异而单独聚为一组; 其他品种聚为一组, 其中‘凤凰木’和‘夕照’的花瓣均具有长的瓣爪、花色橙红, 前者是从实生苗中选育的, 而后者是从前者的扦插苗变异中选育的, 因此, 二者相似性系数最大, 但是一为单瓣花, 一为重瓣花, 从观赏角度而言, 2个品种不宜合并.
可见, 除了目前归于傲大贴梗海棠的2个品种(‘猩红与金黄’、‘碧雪’)外, 同一个种内的品种优先相聚, 具有相似形态特征的品种优先相聚, 品种间的亲缘关系比较清晰. 而且, 从聚类图上还可看出, 贴梗海棠、日本木瓜和傲大贴梗海棠三者的亲缘关系较近, 而与木瓜海棠的亲缘关系较为疏远.
3 讨论我国木瓜属植物种质资源较为丰富, 但栽培品种的植物学归属存在争议. 传统的经典分类以表型的形态标记为依据, 而分子标记的分类基于基因组DNA多态性检测结果. AFLP分析表明, 相同来源的品种优先聚为一类, 木瓜海棠内品种的多态性带最多, 含有最丰富的遗传多态性位点; 傲大贴梗海棠位于日本木瓜和贴梗海棠之间, 且与后者先聚类, 既验证了傲大贴梗海棠的杂交起源, 也说明它与贴梗海棠的亲缘关系比与日本木瓜的亲缘关系更近.
关于木瓜属观赏品种的分类, 王嘉祥(2004)曾提出按照“种源”(即品种的来源)作为品种分类的前提, 但还是将所有观赏品种均归入了贴梗海棠, 而该文中所记载的品种实际上应归于贴梗海棠、木瓜海棠、日本木瓜和傲大贴梗海棠等不同种类; 张茜等(2005)则将‘红宝石’、‘绿宝石’等品种归入毛叶木瓜, 这些品种的植物学归类均没有得到遗传多样性分析的支持. 本研究基本支持Brickell(1996)、凯特·易斯瑞顿等(2004)等的分类处理, 但比较特别的是‘猩红与金黄’和‘碧雪’2个品种. Brickell(1996)和凯特·易斯瑞顿等(2004)均将‘猩红与金黄’放在傲大贴梗海棠内, 但该品种与贴梗海棠关系密切, 除花色为红色外, 其余特征如花朵较小、单瓣等与贴梗海棠原种的差别很小, 因此与贴梗海棠的其他品种相聚成一类, 结合孢粉学(郑林等, 2008)和数量分类的研究结果, 该品种归入贴梗海棠内为宜. 臧德奎等(2007)基于形态学资料, 亦将‘碧雪’归入傲大贴梗海棠, 但该分类也没有得到分子标记的支持, 该品种的地位值得进一步研究.
黄久香, 黄妃本, 许涵, 等. 2008. 海南岛青梅AFLP标记的遗传多样性. 林业科学, 44(5): 46-52. DOI:10.3321/j.issn:1001-7488.2008.05.010 |
凯特·易斯瑞顿, 丹尼斯·伊沃尔德. 2004.世界园林乔灌木.包志毅, 译.北京: 中国林业出版社, 201-202.
|
沈镝, 朱德蔚, 李锡香. 2005. 云南芋种质资源遗传多样性的AFLP分析. 园艺学报, 32(3): 449-453. DOI:10.3321/j.issn:0513-353X.2005.03.013 |
王嘉祥. 2004. 山东省皱皮木瓜品种分类探讨. 园艺学报, 31(4): 520-521. DOI:10.3321/j.issn:0513-353X.2004.04.023 |
杨朝东, 王健, 张俊卫, 等. 2005. 梅花不同样本间亲缘关系的AFLP初步分析. 中国农业科学, 38(10): 2084-2089. DOI:10.3321/j.issn:0578-1752.2005.10.020 |
俞德浚. 1974.蔷薇科//中国植物志编辑委员会.中国植物志: 第36卷.北京: 科学出版社, 348-354.
|
臧德奎, 王关祥, 郑林, 等. 2007. 我国木瓜属观赏品种的调查与分类. 林业科学, 43(6): 72-75. |
张茜, 王光, 何祯祥. 2005. 木瓜种质资源的植物学归类及管理原则. 植物遗传资源学报, 6(3): 339-343. DOI:10.3969/j.issn.1672-1810.2005.03.021 |
赵红霞. 2003. 观赏木瓜. 落叶果树, (2): 50-51. DOI:10.3969/j.issn.1002-2910.2003.02.024 |
郑林, 陈红, 张雷, 等. 2008. 木瓜属植物的花粉形态及品种分类. 林业科学, 44(5): 53-57. DOI:10.3321/j.issn:1001-7488.2008.05.011 |
Aggarwal R K, Brar D S, Nandi S, et al. 1999. Phylogenetic relationship among Oryza species revealed by AFLP markers. Theoretical and Applied Genetics, 98: 1320-1328. DOI:10.1007/s001220051198 |
Bohn M, Utz H F, Melchinger A E. 1999. Genetics similarities among winter wheat cultivars determined on the basis on RFLPs, AFLPs, and SSRs and their use for predicting progeny variance. Crop Sci, 39: 228-237. DOI:10.2135/cropsci1999.0011183X003900010035x |
Brickell C. 1996. Encyclopedia of Garden Plants. London, New York, Stuttgart, Moscow: Dorling Kindersley, 250-252.
|
Dong N V, Subudhi P K, Luong P N, et al. 2000. Molecular mapping of a rice gene conditioning thermosensitive genic male sterility using AFLP, RFLP and SSR techniques. Theoretical and Applied Genetics, 100: 727-734. DOI:10.1007/s001220051345 |
Fang G S H, Grumet R. 1992. A quick and inexpensive method for removing polysaccharides from plant genomic DNA. Biotechniques, 13: 52-57. |
Tomkins J P, Wood T C, Barnes L S, et al. 2001. Evaluation of genetic variat ion in the daylily (Hemerocallis spp.) using AFLP markers. Theoretical and Applied Genetics, 102: 489-496. DOI:10.1007/s001220051672 |