 2009, Vol. 45
2009, Vol. 45文章信息
- 万猛, 田大伦, 樊巍, 李庆云.
- Wan Meng, Tian Dalun, Fan Wei, Li Qingyun
- 豫东平原杨农复合系统物质生产与碳截存
- Biomass Production and Carbon Sequestration in Poplar-Crop Agroforestry Ecosystems in Eastern Henan Plain
- 林业科学, 2009, 45(8): 27-33.
- Scientia Silvae Sinicae, 2009, 45(8): 27-33.
-
文章历史
- 收稿日期:2008-12-25
-
作者相关文章



2. 中南林业科技大学 长沙 410004;
3. 河南省林业科学研究院 郑州 450008;
4. 北京林业大学 北京 100083
2. Central South University of Forestry and Technology Changsha 410004;
3. Henan Academy of Forestry Zhengzhou 450008;
4. Beijing Forestry University Beijing 100083
农林复合系统由于林分、农作物和土壤之间的相互作用而对储藏和固定碳具有巨大的潜力(Nair, 1998). 它具有在林分和土壤中直接固定碳的能力, 其能够通过休耕和禁止滥伐森林来抵消大气中的温室气体(Dixon, 1995; Nair et al., 2002). Dixon等(1993)曾经作过大量研究, 估计每建立1 km2的农林复合经营模式, 就相当于阻止了5 km2的滥伐森林. Sánchez(2000)的研究也表明把森林变成不同类型的农林复合系统损失的碳比转换成耕地或者牧场损失的碳少. 因此, 通过大量的研究表明农林复合系统已经成为一种减少大气CO2浓度, 解决全球气候变暖问题的重要土地利用方式之一. 在未来10年或者1个世纪内, 随着世界上农林复合经营区域大面积的增加, 其将会对陆地生物圈中通量的输入和碳的长期贮藏产生重要的影响(Dixon, 1995). 农林复合系统对碳固定和减少大气中CO2浓度的重要作用正在被人们广泛认同. 但是, 许多学者对农林复合系统研究的焦点多集中在农林复合系统土壤的各种效益、水源涵养、作物和牧场生产力、水分和光照的竞争以及系统的结构配置、养分循环等方面上. 对整个农林复合系统生物量、生产力以及从森林或者单一作物系统向农林复合系统转换时, 有关农林复合系统林分各部分的碳积累率和农林复合系统碳截存等方面的定量分析数据还非常缺乏(Oelbermann et al., 2004). 虽然也有一些关于杨树(Populus)人工林生态系统生物量、生产力、碳贮量以及碳平衡等方面的研究, 但都仅仅是对杨树人工林林分的研究. 本文主要应用标准样地、回归分析等方法, 对豫东平原不同年龄段的杨农间作复合系统的生物量、生产力以及碳截存量进行了研究, 在丰富基础资料的同时, 也为进一步准确评估豫东平原农区农林复合系统的物质生产、碳截存量、农林复合模式的生态效益和经济效益提供了科学的依据.
1 研究区概况研究区位于河南省商丘市民权国营林场, 该林场位于豫东平原黄河故道腹地民权县境内, 地理坐标为34.31°—34.52° N, 114°—115°28′ E, 海拔60.90 m, 年均气温14.0 ℃, 绝对最低气温-16.0 ℃, 大于10 ℃积温4 700 ℃, 无霜期213 d, 年均降水量679 mm, 属于暖温带大陆性季风气候区. 研究区土壤为黄河冲击而形成的风沙土, pH值8.85~9.28, 地下水位1.50~4.00 m.
调查了杨农间作复合系统中杨树5, 9, 11, 13年4个龄级的生物量, 各龄级林分林相比较整齐, 其株行距分别为6 m×9 m, 3 m×10 m, 8 m×30 m, 3 m×30 m, 平均胸径分别为20.30, 24.00, 34.20, 40.00 cm, 平均树高分别为12.20, 22.90, 26.90, 29.60 m, 由于立地条件和栽植密度不同, 各林带杨树生长状态有所不同. 4个龄级的杨农间作复合系统中在夏季和秋季间作的农作物分别是小麦(Triticum aestivum)和花生(Arachis hypogaea).
2 研究方法 2.1 标准地设置样地调查于2007年7—8月进行, 按照林带株行距的不同, 研究区内设置标准样地22块, 标准地面积根据林带株行距、杨树栽植密度而定, 4个龄级的农林复合间作样地面积分别设置为12 m×54 m, 20 m×33 m, 64 m×33 m, 64 m×33 m, 每个样地都包含4行林带, 对样地内林木按每木检尺法, 调查林木的胸径(DBH/cm)、高度(H/m)、活枝下高(HB/m)、冠幅(EW/m×SN/m)、树龄(A/a)、郁闭度和株数.
2.2 单木生物量测定采用Monsi(1974)、木村允(1981)的“分层切割法”结合树干解析数据对每个龄级所选取的3株标准木进行生物量测定, 分层测定树干、树枝、树叶和树皮的鲜质量并选取各组分样本. 根部用“分层挖掘法”(0~20, 20~40, 40~60, 60 cm以上)进行挖掘, 测定根系鲜质量并取样. 将所取林木的各器官样品在85 ℃下烘干至恒量, 计算出各器官干物质质量, 由此推算单株各器官生物量及全株生物量. 生物量采用木村允(1981)的基本公式建立回归模型.
本文研究的净生产力指平均净生产力, 由于条件所限未对动物、昆虫等取食量作专门调查, 净生产力计算按木村允(1981)的平均生产力(PNM)是森林总生物量(W)被年龄(A)所除之商表示, 即: PNM=W/A.
2.3 杨农间作复合系统生物量测定杨农间作复合系统中的农作物采用直接收获法计算其生物量. 按照杨农间作复合系统中杨树5, 9, 11和13年4个不同龄级, 每个龄级分别设立3条样带, 共12条样带; 在每个样带中由南向北按照2 m间隔设置1 m×1 m小样方, 共39个小样方. 调查样方内农作物小麦、花生的产量、作物秆生物量以及小麦的千粒质量.
杨农间作复合系统生物量采用林分平均胸径、林分平均树高和林分株数密度3个因子, 结合单木生物量回归模型以及夏季和秋季间作的不同作物对杨农间作复合系统的总生物量进行测算.
2.4 杨农间作复合系统林木凋落物量的测定按照杨农间作复合系统中杨树5, 9, 11和13年4个不同龄级, 分别设立1块40 m×40 m固定标准地. 凋落物的收集是通过简易的凋落物收集器进行收集, 收集器的规格为50 cm×50 cm×60 cm. 在5, 9, 11和13年4个不同龄级的固定样地分别设置凋落物收集器14, 14, 29和29个. 从2007年7月起于每月底收集凋落物, 凋落物收集后带回实验室, 进行分类, 将各组分凋落物分别置于烘箱中, 80 ℃下烘干至恒量, 称量.
2.5 杨农间作复合系统植物碳含量的测定及碳截存量的计算方法选取植物(林木、农作物)各组分的鲜样, 在实验室80 ℃恒温下烘至恒量并粉碎, 过60目网筛, 采用重铬酸钾、硫酸氧化-外加热法(中国土壤学会农业化学专业委员会, 1983)测定植物组分中有机碳含量. 本文研究的碳截存指植物(林木、农作物)的碳截存量, 土壤的碳截存量未计算在内. 根据植物体(林木、农作物)地上、地下生物量和植物(林木、农作物)各组分的碳含量, 使用下式计算碳截存量:

|
式中: C为碳截存量(t·hm-2); M为地上和地下部分生物量; a为植物(林木、农作物)各组分的碳含量的平均值.
3 结果与分析 3.1 单木生物量模型将测定的12株标准木生物量数据代入W=a(D2H)b和W=aDb式进行回归分析, 得出不同林龄杨农间作复合系统中林分各器官及单株的生物量模型(表 1).
|
|
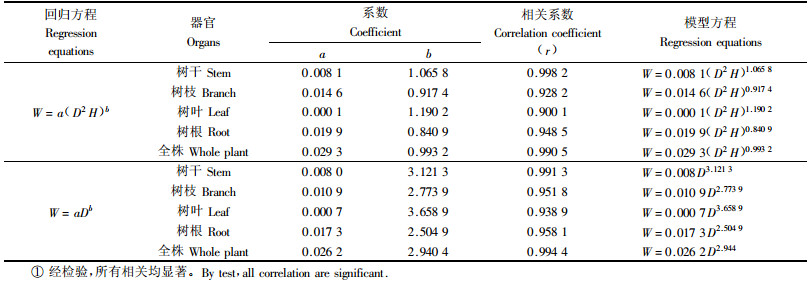
从表 1可以看出: 两式相关系数均很高, 最低的为0.900 1, 最高的为0.998 2, 多数在0.94以上, 尤其是2种形式的全株总生物量回归方程的相关系数均在0.99以上, 表明各器官生物量和D2H和D有良好的相关性, 均可用于计算农林复合系统中林木层的生物量. 经F检验, 各估测模型的相关程度均达到了显著(α=0.05)或极显著水平(α=0.01). 在林业调查应用中具有较高的参考价值. 从回归模型看, 采用W=a(D2H)b对树干进行估计比采用W=aDb对树干进行估计相关系数高, 但在实际推广应用中, 由于杨树生长迅速, 13年生的杨树高可达34 m, 树高的测定就相对困难, 误差也比较大, 因此, 对各组分和全株的估测模型采用以胸径为自变量的回归方程W=aDb, 其实用性更强(郑景明等, 1998).
3.2 杨农间作复合系统中林分生物量 3.2.1 杨农间作复合系统不同林龄林分生物量豫东平原杨农间作复合系统中林分生物量调查结果及分布特点如表 2所示, 从表 2中可以得出5, 9, 11和13年杨农间作复合系统林分总生物量分别为16.35, 83.81, 99.50和127.82 t·hm-2. 其中, 4个龄级杨农间作复合系统林分单株的各组分生物量是树干所占的比例最大, 分别为47.00%, 60.70%, 57.80%和59.00%, 说明生物量的积累主要是树干生物量的增加. 从表 2中看出, 树干、树枝、树叶和树根各器官生物量都具有不同程度的增长, 总的趋势是随着林龄的增长而逐渐积累增多, 而相对生长速率则逐渐减少. 另外可以看出11年生林分单株各组分的生物量与9年生林分单株各组分的生物量差别不大, 这是由于9年生林分的栽植密度较大, 所以其总生物量与11年生林分的总生物量接近, 因此也就造成了其树干生物量与11年生杨树树干生物量相差不大.
|
|
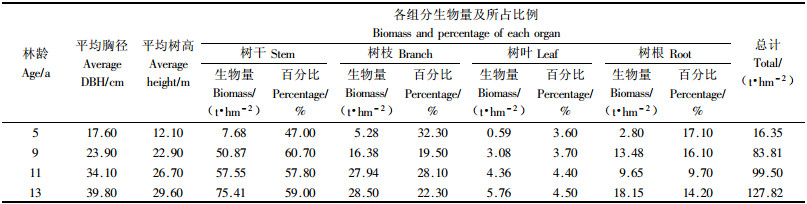
从表 2还可以得出, 随着林龄的增长各年龄段单株根系生物量迅速增加, 可占其林分生物量的10%~20%, 而粗根生物量是根系生物量的主要组成部分(Fogel, 1983), 根据研究估算得出粗根、中根、小根分别占根系生物量的87.30%, 10.20%, 2.50%, 并且随着林龄的增加粗根所占根系生物量比例也有所增加. 地下部分中的粗根和中根生物量占根系总生物量的大部分, 达到了97.50%, 起着良好的机械支持作用. 吸收根(小根和细根)生物量虽然仅占根系总生物量的较小比例, 但在森林生态系统中起着吸收水分与养分的作用(杨玉盛等, 2003).
5, 9, 11和13年杨农间作复合系统林分单株生物量的分配格局呈现出: 树干>树枝>树根>树叶的分配格局, 树干占的比例最大, 可达到47.00%~63.60%, 其次是树枝、树根, 树叶生物量所占比例最小. 说明树干、树枝的干质量占该林分单株总干质量比例大而且增长趋势快, 这也是总干质量主要的积累过程, 树干、树枝更好地起着支持、输导和吸收作用, 以维持林分在生存空间的持续生长.
杨树属于速生树种, 从5年生杨树到13年生杨树其总生物量从16.35 t·hm-2增加到127.82 t·hm-2, 随着林龄的递增林分的生物量增长速度也在递增(图 1), 其中树干生物量增加了67.73 t·hm-2, 枝生物量增加了23.22 t·hm-2, 叶生物量增加了5.17 t·hm-2. 13年生的杨树林分生物量可达127.82 t·hm-2, 这也充分说明了杨树速生、经济效益高的优点. 从图 1可以看出9年生林分的生物量至11年生林分的生物量变化趋于平缓, 这是由于2个年龄段林分的栽植密度不同造成的. 在样方调查中, 11年生杨树平均胸径为34.10 cm, 最大胸径可达到40.70 cm, 林带内间作小麦、花生等农作物, 所以株行距相对大些, 为8 m×30 m, 栽植密度为105株·hm-2, 而9年生的杨树林分株行距为3 m×10 m, 栽植密度达到303株·hm-2, 所以造成这2个林龄的杨树林分总生物量相差不大的现象. 由此可以得出林分的生物量与林分年龄、栽植密度、立地条件等有很大关系.

|
图 1 林分生物量与林龄之间的曲线关系 Figure 1 The relation between stand biomass and its age |
凋落物数量组成及质量是林木生长发育过程中的新陈代谢产物, 受林木组成结构等影响, 其是森林自我培肥、恢复地力主要途径之一(杨玉盛, 1998). 经测定, 5, 9, 11和13年4个不同林龄农林复合系统年凋落物总量分别为11.20, 12.90, 13.80和13.90 t·hm-2. 4个不同林龄农林复合生态系统年凋落物量随着林龄的增大而增大. 年凋落物量的变化除了与所处不同气候带有关外, 主要与林龄差异有关, 因此林龄的差异决定了这4种类型年凋落物总量的变化.
3.3 杨农间作复合系统中农作物生物量豫东平原农区杨农间作复合系统在夏季和秋季间作的农作物是小麦和花生. 杨农间作复合系统中杨树5, 9, 11和13年4个龄级的株行距分别为6 m×9 m, 3 m×10 m, 8 m×30 m, 3 m×30 m. 经测算, 因林龄、树高、冠幅和株行距的不同间作的农作物产量和生物量也不同. 小麦是豫东平原农区最重要的夏季作物. 在4个龄级中间作的小麦总生物量分别是5.88, 6.95, 9.80和10.42 t·hm-2(表 3), 其中籽实分别占总生物量的46.30%, 43.90%, 53.40%和51.20%; 茎叶分别占总生物量的35.20%, 33.70%, 30.10%和31.80%; 根分别占总生物量的18.50%, 22.50%, 16.50%和17.10%. 秋季作物花生在4个龄级中间作的总生物量分别是10.80, 12.30, 15.00和16.80 t·hm-2(表 3), 其中籽实分别占总生物量的35.65%, 34.55%, 37.80%和38.00%; 茎叶分别占总生物量的62.80%, 57.60%, 52.40%和53.40%; 根分别占总生物量的1.70%, 8.00%, 9.50%和11.20%.
|
|
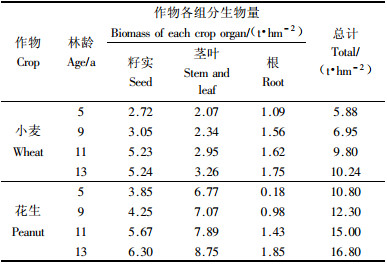
豫东平原农区5, 9, 11和13年4个龄级杨农间作复合系统农作物一季的总生物量分别是16.68, 19.25, 24.80和27.04 t·hm-2. 对4种不同经营模式的小麦和花生总生物量以及各个组分生物量的比较结果表明, 宽行的生物量高于窄行的生物量, 此外, 农作物生物量还受到林木树高、冠幅以及病虫害的影响.
3.4 杨农间作复合系统生物量通过上述计算分析可以得到豫东平原农区5, 9, 11和13年杨农间作复合系统总生物量分别为33.03, 103.06, 124.30和154.60 t·hm-2.
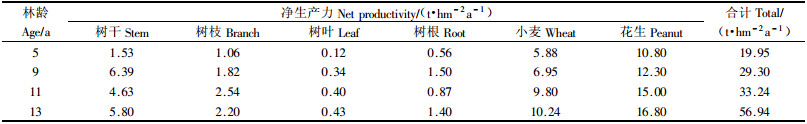
3.5 杨农间作复合系统净生产力测算由于研究区属于杨农间作复合系统, 林带内间作花生、小麦等农作物, 林下无灌木, 仅在林带边缘有极少量草本植物, 故本文杨农间作复合系统生产力指的是林分的净生产力和间作农作物的净生产力之和, 测定结果见表 4. 从表 4看出, 这4个年龄段杨农间作复合系统的总净生产力分别为19.95, 29.30, 33.24和56.94 t·hm-2a-1, 都具有较高的生产力. 其中4个年龄段杨农间作复合系统林分的净生产力分别为3.27, 10.05, 8.44和9.83 t·hm-2a-1, 特别是杨农间作复合系统中从9年生到13年生杨树的生产力水平很高, 说明这一阶段杨树生长速度很高. 调查中还发现树干的净生产力最大, 分别占林分净生产力的46.79%, 63.58%, 54.86%和59.00%, 其次是树枝、树根, 树叶净生产力所占比例最小. 在计算过程中未考虑枯损量及昆虫等取食量, 故林分净生产力可能要比实际净生产力偏小(梁妮等, 2006). 4个年龄段杨农间作复合系统作物的净生产力分别为16.68, 19.25, 24.80和27.04 t·hm-2a-1.
|
|
从表 5可以得出, 杨树各器官含碳率分别是: 树干0.452 7, 树枝0.457 4, 树叶0.448 2, 树根0.458 7, 各器官之间含碳率变异系数仅为2%, 平均含碳率为0.450 7, 凋落物平均含碳率为0.435 9. 器官中树根的碳含量最大, 树叶的碳含量最小, 表现出树根>树枝>树干>树叶>凋落物的分配格局. 杨农间作复合系统在夏季和秋季间作的农作物小麦和花生的平均含碳率分别是0.458 3和0.45.
|
|

从表 6可以得出豫东平原杨农间作复合系统5, 9, 11和13年杨农间作碳截存量分别为19.86, 52.42, 62.45和76.44 t·hm-2. 其中, 林分(包括凋落物)的碳截存量分别为12.31, 43.69, 51.21和64.11 t·hm-2, 林分的碳截存量占杨农间作复合系统总碳截存量的比率是61.98%~83.87%. 碳截存量随着林龄的增加而增加, 除5年杨农间作碳截存量比较小外, 其他3个年龄段的杨农间作碳截存量逐步递增. 同龄不同类型的植被碳截存量不同, 同一类型植被不同生长阶段碳截存量都差异很大, 杨树作为一种速生树种, 生物量每年都会有很大差异, 所以不同地区不同年龄段杨树的碳截存量是不同的. 4个年龄段, 树干的碳截存量最大, 其次是树枝、树根和树叶.
|
|
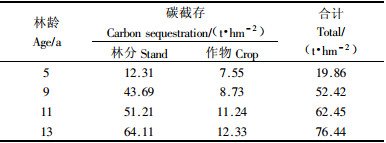
豫东平原杨农间作复合系统5, 9, 11和13年杨农间作作物碳截存量分别为7.55, 8.73, 11.24和12.33 t·hm-2. 作物的碳截存量占杨农间作复合系统总碳截存量的比率是16.13%~38.02%. 夏季作物小麦的碳截存量低于秋季作物花生的碳截存量, 平均低于其5.50%, 说明花生具有较高的固碳能力. 但是, 如果对小麦秸秆、花生秧处理不当, 如直接焚烧、不实行轮作和免耕等, 就会引起大量的CO2的排放和流失, 所以应当采取适当措施, 如秸秆还田、保留残茬等, 不但可以增加农田中土壤的有机质含量, 并能有效增加土壤肥力, 代替部分化肥, 减少化肥投入量, 具有良好的生态效益和经济效益.
4 结论与讨论利用生长关系分别建立了豫东平原农区5, 9, 11, 13年4个不同年龄段的农林复合杨树人工林单木及各个器官生物量的2种回归模型, 即W=a(D2H)b和W=aDb, 其相关程度均达到显著水平, 尤其是2种形式的总生物量回归方程的相关系数均在0.99以上, 表明有良好的相关性, 可以在生产实践中推广应用.
豫东平原农区5, 9, 11和13年杨农间作复合系统生物量分别为33.03, 103.06, 124.3和154.6 t·hm-2. 与查同刚(2007)所研究的北京大兴6年生杨树人工林生物量25.2 t·hm-2, 8年生杨树人工林的生物量53.10 t·hm-2相比. 此次调查的5年生杨农间作林分生物量略低, 9年生杨农间作林分生物量明显较高. 而与Liang等(2006)对山东9年生杨树人工林生物量的研究结果(平均139.00 t·hm-2)相比明显低. 这与杨农间作复合系统和杨树人工林生态系统的栽植密度和经营模式侧重点不同有关. 可见, 合理的间作年限和合理的间作行距, 不但可以有效地提高作物产量, 而且可以提高林木的生长量和培养大径材, 取得最佳的经济效益和生态效益(Imo et al., 1999).
豫东平原农区5, 9, 11和13年杨农间作复合系统总净生产力分别为19.95, 29.30, 33.24和56.94 t·hm-2a-1. Lodhiyal等(1994)研究4年生杨树人工林高密度栽植(666株·hm-2)的净生产力是32.40 t·hm-2a-1, 5年生杨树人工林(栽植密度400株·hm-2)的净生产力是20.00 t·hm-2a-1, 高于豫东平原农区5年杨农间作复合系统(栽植密度155株·hm-2)的净生产力. 但是2地区的栽植密度不同, 其栽植密度远远大于豫东平原杨农间作复合系统的栽植密度. Liang等(2006)对山东9年生杨树人工林的净生产力的研究结果是15.42 t·hm-2a-1, 远低于9年杨农间作复合系统的净生产力. 与国内其他杨树人工林同类的研究相比, 豫东平原农区9年杨农间作复合系统的净生产力还高于西藏、新疆等西北水分匮乏地区的9年生杨树人工林的净生产力.
豫东平原农区5, 9, 11和13年杨农间作复合系统碳截存量分别为19.86, 52.42, 62.45和76.44 t·hm-2. 林分和作物是农林复合系统的重要组成部分(Livesley et al., 2000). 不同年龄段的林分对农林复合系统碳截存的贡献率不同. 林分在整个生态系统的构建中起着极其重要的作用, 5年生的林分正处于快速生长期, 其碳截存作用正在逐步加强, 其碳截存潜力仍然非常巨大. 农作物的碳截存作用存在比较大的争议. 如果仅从碳生产的角度看, 农作物对整个生态系统的贡献也比较大, 4个年龄段农作物碳截存分别占总碳截存量的38.02%, 16.65%, 18.00%和16.13%. 陈军(2007)研究的北京大兴6年生杨树人工林碳储量是26.33 t·hm-2. 唐罗忠等(2004)分析了江苏里下河地区10年生杨树人工林的碳储量动态, 结果显示人工林碳储量为74.10 t·hm-2. 通过比较分析得出本试验区杨农间作复合系统林分碳截存量都低于相应年龄段的杨树人工纯林的碳储量, 其原因在于: 杨树人工林的栽植密度明显要高于本研究区杨农间作复合系统的栽植密度. 杨树人工纯林林下植被茂密, 凋落层较厚, 固定了大量的碳. 本试验区是风沙土, 土质疏松通透性好, 保肥蓄水能力差, 杨农间作复合系统中林分整个生长季大部分时间处于水分胁迫状态, 导致叶片净光合速率降低, 这必然影响其碳截存的能力. 但是, 实施农林复合经营可以改变农田土壤理化性质, 提高土壤肥力, 使土壤的固碳能力得到提高.
陈军.2007.杨树人工林地上生物量和碳储量研究.北京林业大学博士学位论文, 41-44. http://cdmd.cnki.com.cn/Article/CDMD-10022-2007076907.htm
|
梁妮, 王卫斌, 田昆. 2006. 4年生及13年生西南桦人工林生物量的分布特征. 西部林业科学, 35(4): 118-122. DOI:10.3969/j.issn.1672-8246.2006.04.022 |
Monsi M.1974.植物群落的数学模型.植物生态学译丛(第一集).北京: 科学出版社, 123 -144.
|
木村允. 1981. 陆地植物群落生产量测定法. 北京: 科学出版社, 70-71.
|
唐罗忠, 生原喜久雄, 黄宝龙, 等. 2004. 江苏省里下河地区杨树人工林的碳储量及其动态. 南京林业大学学报, 28(2): 1-6. DOI:10.3969/j.issn.1000-2006.2004.02.001 |
杨玉盛, 陈光水. 2003. 格氏栲天然林与人工林细根生物量、季节动态及净生产力. 生态学报, 23(9): 1719-1730. DOI:10.3321/j.issn:1000-0933.2003.09.004 |
杨玉盛. 1998. 杉木林可持续经营的研究. 北京: 中国林业出版社, 32-40.
|
查同刚.2007.北京大兴杨树人工林生态系统碳平衡的研究.北京林业大学博士学位论文, 45-58. http://cdmd.cnki.com.cn/Article/CDMD-10022-2007077364.htm
|
郑景明, 张育红. 1998. 林分生物量研究综述. 辽宁林业科技, (4): 43-45. |
中国土壤学会农业化学专业委员会. 1983. 土壤农业化学常规分析方法. 北京: 科学出版社, 272-273.
|
Dixon R K. 1995. Agroforestry systems: sources or sinks of greenhouse gases. Agroforestry Systems, 31: 99-116. DOI:10.1007/BF00711719 |
Dixon R K, Winjum J K, Schroeder P E. 1993. Conservation and sequestration of carbon: the potential of forest and agroforestry management practices. Global Environmental Change, 3: 159-173. DOI:10.1016/0959-3780(93)90004-5 |
Fogel R. 1983. Root turnover and productivity of coniferous forests. Plant and Soil, 71: 75-85. DOI:10.1007/BF02182643 |
Imo M, Timmer V R. 1999. Vector competition analysis of a Leucaena-maize alley cropping system in western Kenya. Forest Ecology and Management, 47: 1-14. |
Lodhiyal L S, Singh R P, Singh S P. 1994. Productivity and nutrient cycling in popular stands in central Himalaya, India. Canadian Journal of Forest Research, 24: 1199-1209. DOI:10.1139/x94-158 |
Liang W J, Hu H Q, Liu F J, et al. 2006. Research advance of biomass and carbon storage of poplar in China. Journal of Forestry Research, 17(1): 75-79. DOI:10.1007/s11676-006-0018-0 |
Livesley S J, Gregory P J, Buresch R J. 2000. Competition in tree row agroforestry systems: 1. Distribution and dynamics of fine root length and biomass. Plant Soil, 227: 149-161. DOI:10.1023/A:1026551616754 |
Nair R P K. 1998. Directions in tropical agroforestry research: past, present, and future. Agroforestry Systems, 38: 223-245. |
Nair R P K, Nair V D. 2002. Carbon sequestration in agroforestry systems//Proceedings of the 17th World Congress on Soil Science, 2002-08-14-18, Thailand. https://link.springer.com/chapter/10.1007/978-94-007-7723-1_10
|
Oelbermann M, Voroney R P, Gordon A M. 2004. Carbon sequestration in tropical and temperate agroforestry systems: a review with examples from Costa Rica and southern Canada. Agriculture Ecosystems and Environment, 104: 359-377. DOI:10.1016/j.agee.2004.04.001 |
Sánchez P A. 2000. Linking climate change research with food security and poverty reduction in the tropics. Agriculture Ecosystems and Environment, 82: 371-383. DOI:10.1016/S0167-8809(00)00238-3 |