 2009, Vol. 45
2009, Vol. 45文章信息
- 赵仲辉, 康文星, 田大伦, 项文化, 闫文德.
- Zhao Zhonghui, Kang Wenxing, Tian Dalun, Xiang Wenhua, Yan Wende
- 湖南会同杉木液流变化及其与环境因子的关系
- Sap Flow Rate and Its Relationship with Environmental Factors of Chinese Fir Plantation in Huitong, Hunan Province
- 林业科学, 2009, 45(7): 127-132.
- Scientia Silvae Sinicae, 2009, 45(7): 127-132.
-
文章历史
- 收稿日期:2008-03-16
-
作者相关文章


在土壤-植物-大气连续体构成的水流路径中,树干液流量制约着整个树冠的蒸腾量,如何直观、精确和连续测定水分在木质部中的流动速率是科学家们一直在探索和渴望解决的问题(Baker et al., 1987)。近几十年来,国内外相关领域的学者们相继提出了一些诸如称重法、截干法和茎流速率法等不同的测定方法,目前茎流速率法应用最为广泛。茎流速率法主要包括放射法同位素示踪法、染色法、热脉冲法、热扩散探针法、热平衡法和核磁共振图像扫描法,这些方法在使用上都有一定的适用性和局限性(刘奉觉等,1997)。热技术方法具有时间分辨率高、对植物破坏小、野外操作方便及连续自动监测等优点,20世纪90年代以来,我国学者应用该技术对核桃楸(Juglans mandshurica)(严昌荣等,1999)、白桦(Betula platyphylla)(孙慧珍等,2002;2005)、桉树(Eucalyptus spp.)(尹光彩等,2003)、白杨(Populus tomentosa)(常学向等,2004)、油松(Pinus tabulaeformis)、栓皮栎(Quercus variabilis)(聂立水等,2005)、马占相思(Acacia mangium)(马玲等,2005)、银杏(Ginkgobiloba)(孙守家,2006;刘健等,2007)、侧柏(Platycladus orientalis)(王华田,2006)、胡杨(Populus euphratica)(张小由等,2006;白云岗等,2007;司建华等,2007)、麻栎(Quercus acutissima)(赵文飞等,2007)、龙爪槐(Sophora japonica)(谢恒星等,2007)和尖峰栲(Castanopsis jianfenglingensis)(张刚华等,2007)等树种的液流速率进行了研究,并分析了液流与环境因子的关系,通过适当的尺度转换还可研究单木或林地的蒸腾耗水状况(马李一等,2001;熊伟等,2003;张小由等,2006;赵平等,2006;Bovard et al., 2005;Gazal et al., 2006)。
本文应用ICT2000TE自动监测系统对杉木(Cunninghamia lanceolata)树干液流速率和环境因子进行同步观测,研究杉木树干液流变化规律及其与主要环境因子之间的关系,为尺度转换计算单木或林分蒸腾量和建立杉木耗水机理模型提供基础数据和科学依据。
1 试验地概况试验地位于湖南会同杉木林生态系统国家野外科学观测研究站(109°45′E,26°50′N)。地貌为低山丘陵,海拔270~400 m;地层属震旦纪板溪系灰绿色变质板页岩, 土壤为山地黄壤,土层深厚,土壤肥力条件好;该区年均气温16.8℃,1月平均气温4.4℃,7月平均气温26.3℃;年均相对湿度80%,年降水量1 100~1 500 mm,降水主要集中于4—8月,属典型的亚热带湿润季风气候。
试验在该站第Ⅲ集水区进行,集水区面积19 483.0 m2,主流长230.0 m,平均宽度84.7 m,流域形状指数0.37,平均坡度25.0°。林分为1988年春季全垦整地后营造的第2代杉木人工林,造林后前3年(1988—1990年)每年的7和11月各全垦抚育1次。林相整齐,长势良好,林分密度2025株·hm-2,平均胸径为14.6 cm,平均高14.2 m(2005-11)。
2 研究方法根据2块面积各为666.7 m2的标准样地的每木检尺(2005-11)结果,选择干形通直、冠形良好的平均木1株,样木树高为13.0 m,胸径为13.7 cm,试验时间为2005-12—2006-11。2006-12,利用生长锥钻取木芯,确定样木边材面积为71.2 cm2。
2.1 树干液流的测定树干液流用热扩散方法(Granier, 1985;1987)进行测定。热扩散探头(Dynamax, Houston TX, USA)由2根探针组成,上部探针恒定连续加热,内有加热线和热电偶;下部探针为参考端。通过测定2根探针在边材的温差值,可由Granier的经验公式(Granier, 1985)求出树干边材的液流速度或液流密度的连续变化:
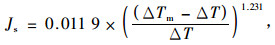
|
式中:Js为液流速率(cm·s-1);ΔTm为无液流时两探针最大温差(℃);ΔT为两探针测定的温差(℃)。
2.2 环境因子的测定利用观测铁塔测定林内1.5 m高处的空气温度(℃)和相对湿度(%)(RH1,Vaisala,UT,USA)、林冠下空隙中的太阳总辐射(W·m-2)(SKS1110E Silicon Cell Pyranimeter, Skye, UK)和光合有效辐射(μmol·m-2s-1) (SKP215 PAR Quantum Sensor, Skye, UK),同步测定土壤温度(℃)(20 cm深处) (Delta-T Devices Ltd., Cambridge, UK)和土壤水分含量(%)(20,30和50 cm深处)(ECH2O, Decagon Devices, Inc., USA)。以上林内环境因子探头与热扩散探头通过ZENO3200 (Coastal Environmental Systems, USA)数据采集器控制,每5 s采集1次数据,每15 min取平均值并记录1次数据。在铁塔的顶部用常规气象仪器测定冠上空气温度(℃)和冠上相对湿度(%),并通过计算得到冠上的水汽压(hPa)和饱和差(hPa)。2006年7月,在生态站内的气象观测场(距测点直线距离约250 m)中安装有自动气象站,进行林外空气温度(℃)、相对湿度(%)、风速(m·s-1)、太阳总辐射(W·m-2)和降水量(mm)等气象要素的测定。
2.3 数据处理假定树干边材中液流速率处处相等,则单木液流速率Tr(g·s-1)可由下式计算:
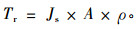
|
式中:A为边材面积(cm2), ρ为水的密度(g·cm-3)。单木日液流量Tvd(kg·d-1)由下式计算:
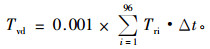
|
式中:Δt为采样时间间隔(s),在本研究中为900 s;Tri为第i时刻的单木液流速率(g·s-1)。单木月液流量Tvm(kg)为

|
式中:n为每月的天数;Tvdj为该月第j天的单木日液流量(kg·d-1)。
数据处理在Excel(Microsoft, USA)和SPSS13.0(SPSS Inc., USA)中完成。
3 结果与分析 3.1 不同季节不同天气单木液流速率的平均日变化不同季节不同天气(晴天和阴天)单木液流速率的平均日变化见图 1。在晴天,冬、春、夏、秋季液流速率开始升高的时间分别出现在09:15,07:15,06:30和07:30前后,峰值分别出现在午后13:00(0.044 8g·s-1),12:30(0.2005g·s-1),12:30(0.3791 g·s-1)和12:15(0.1257 g·s-1)前后,回到低值水平的时间分别为17:00,21:00,22:00和21:15前后;阴天与晴天类似,只是液流速率开始升高的时间相对较晚,回到低值的时间较早,峰值大为降低,冬、春、夏、秋季液流速率开始升高的时间分别在10:30,07:30,07:00和07:45前后,峰值分别出现在午后14:15(0.0074 g·s-1),14:15(0.0680 g·s-1),13:30(0.1683 g·s-1)和12:45(0.0784 g·s-1)前后,回到低值水平的时间分别为18:00,20:00,20:00和19:30前后。

|
图 1 四季晴天和阴天平均液流速率的日变化 Figure 1 Daily variations of mean sapflow rate in sunny and overcast day in four seasons |
晴天,由冬季至夏季,液流开始升高的时刻逐渐提前,回到低值的时刻逐渐推后,由夏季至冬季,液流开始升高的时刻逐渐推后,回到低值的时刻逐渐提前,说明液流的持续时间与昼长变化规律一致。但平均液流启动时间表现为冬季比夏季滞后2.75 h,回到低值时间表现为冬季比夏季提前3.50 h,即冬季液流持续时间比夏季缩短了6.25 h。秋季液流峰值出现的时间最早(12:15),冬季最晚(13:00),但差距不大,相差45 min左右,冬季气温对液流峰值出现时间起主要作用,秋季土壤水分含量减小可能使液流峰值出现时间提早。
3.2 单木日液流量的年变化根据2005-12—2006-11杉木树干液流连续监测的数据,计算出杉木单木日液流量的年变化动态(图 2)。由图 2可见,11月16日至次年2月12日,是杉木的非生长季节,日液流量数值小,在(0.32±0.26) kg·d-1的低值水平小幅波动,1月29日最大(1.62 kg·d-1),2月6日最小(0.04 kg·d-1),低温可能是这一时期液流量小的主要限制因子;2月13日至6月7日,日液流量在(3.22±2.94) kg·d-1变化,3月10日最大(18.19 kg·d-1),3月12日最小(0.12 kg·d-1),变化幅度为18.07 kg·d-1,该时期会同站阴雨天气较多,土壤水分含量多,辐射和气温的日际变化大,从而造成日液流量的日际变化大;6月8日至9月4日,日液流量呈逐波下降的趋势,平均为(7.31±3.95) kg·d-1,6月8日和14日最大(19.50 kg·d-1),8月4日最小(0.57 kg·d-1),变化幅度为18.93 kg·d-1,该时期为1年中辐射最强、温度最高的季节,杉木的蒸腾潜力大,只要有充分的水分供应,液流量可以达到很高的数值,日液流量最大值都出现在6月雨后的晴天;9月5日至11月15日,日液流量维持在(2.18±1.10) kg·d-1的水平,9月22日最大(3.95 kg·d-1),10月8日最小(0.14 kg·d-1),变化幅度为3.81 kg·d-1,该时期降水少,土壤水分含量低,是液流速率降低的主要原因。

|
图 2 2005-12—2006-11日液流量年变化 Figure 2 Yearly change of daily sap flow value from December in 2005 to November in 2006 |
单木月液流量(图 3)表现为1月份最小,只有9.21 kg,6月最大,达277.28 kg。冬季(2005-12—2006-02)、春季(2006-03—2006-05)、夏季(2006-06—2006-08)和秋季(2006-09—2006-11)单木液流量分别为48.84,324.40,651.02和182.63 kg,单木全年的液流量为1 206.89 kg。

|
图 3 月液流量的年变化 Figure 3 Annual changes of monthly sap flow value |
1) 单木液流速率与环境因子的关系以液流速率为因变量,以太阳总辐射、气温、风速、土温、相对湿度和土壤水分含量为自变量,通过逐步线性回归分析,得如下回归方程:
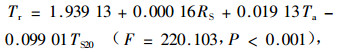
|
式中:RS为太阳总辐射(W·m-2);Ta为空气温度(℃);TS20为20 cm深处的土壤温度。
回归分析表明液流速率与太阳总辐射和气温为正相关关系,与20 cm深处的土温为负相关关系(土温位相滞后效应)。
2) 单木日液流量与环境因子的关系表 1列出了单木日液流量与主要环境因子间的Pearson相关系数。由表 1可见,在不同季节,单木日液流量与各环境因子的相关系数相差较大。从全年来看,单木日液流量与林内总辐射、林内光合有效辐射、冠上气温、林内20 cm深处土温、冠上水汽压、冠上饱和差和50 cm深处土壤水分含量间存在极显著的正相关(P<0.01),与林内30 cm深处的土壤水分含量呈显著的正相关(P<0.05),与冠上空气相对湿度存在极显著的负相关(P<0.01)。冬季单木日液流量与林内总辐射、林内光合有效辐射、冠上气温、冠上水汽压、50 cm深处土壤水分含量间存在极显著的正相关(P<0.01),与林内20和30cm深处的土壤水分含量呈显著正相关(P<0.05)。春季单木日液流量与林内总辐射、林内光合有效辐射、冠上气温、冠上水汽压、冠上饱和差和林内20cm深处土温间存在极显著的正相关(P<0.01),与冠上空气相对湿度存在极显著的负相关(P<0.01)。夏季单木日液流量与林内总辐射、林内光合有效辐射、土壤水分含量(20,30和50 cm深处)及冠上水汽压间存在极显著的正相关(P<0.01),与林内20cm深处土温呈显著负相关(P<0.05),与土壤冠上空气温度呈显著正相关(P<0.05)。秋季单木日液流量与林内总辐射、林内光合有效辐射、林内20cm深处的土壤温度、冠上气温、冠上水汽压、冠上饱和差间存在极显著的正相关(P<0.01),与冠上空气相对湿度、冠上降水量、林内20和30 cm深处的土壤水分含量间存在极显著负相关(P<0.01)。上述环境因子中,有些因子间具有明显的相关性,如冠上气温、冠上饱和差、冠上水汽压和冠上相对湿度相互间及林内太阳总辐射与林内光合有效辐射之间,为消除不同因子间的干扰,以单木日液流量为因变量,以林内太阳总辐射、林内土壤温度、土壤水分含量(20、30和50 cm深处)、冠上气温、冠上相对湿度、降水量为自变量,进行全年、春季、夏季、秋季和冬季的逐步线性回归分析(引入因子P<0.05,剔除因子P<0.10)。
|
|
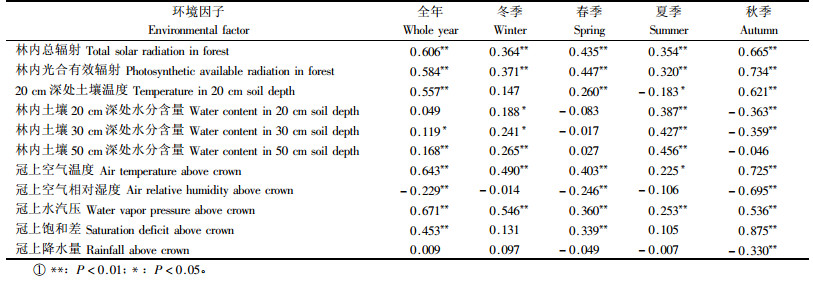
年回归方程:

|
式中:Tau为冠上空气温度(℃);CSW30为30 cm深处的土壤水分含量(%);Sm为林内太阳总辐射(MJ·m-2d-1);CSW20为20 cm深处的土壤水分含量(%)。
冬季回归方程:
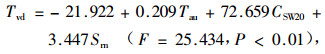
|
春季回归方程:
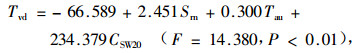
|
夏季回归方程:
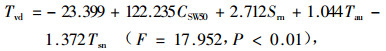
|
式中:CSW50为50 cm深处的土壤水分含量(%);Tsn为林内土壤温度(℃)。
秋季回归方程:
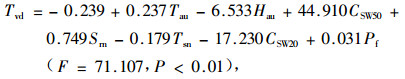
|
式中:Pf为降水量(mm)。
偏相关系数的大小反映了该因子在回归方程中的重要程度,在以上的回归方程中,冠上气温偏相关系数最大,其次是土壤水分含量,再次就是林内总辐射。
3) 单木月液流量与环境因子的关系以单木月液流量为因变量,以20,30和50 cm深处土壤水分含量月均值,冠上气温、冠上相对湿度和林内土壤温度的月均值及林内总辐射和降水量的月总量为自变量,进行逐步线性回归分析,结果如下:

|
式中:Tm为冠上月平均气温(℃);Hm为冠上月平均相对湿度(%)。
4 结论与讨论晴天和阴天,杉木液流速率日变化均呈现为单峰型。杉木的光合速率在自然条件下呈现为单峰型的日变化,没有“光合午休”现象,液流速率与光合速率的变化是完全一致的(肖文发等,2002)。
太阳辐射是光合作用的驱动力,辐射的强弱决定了气孔的开张程度,从而影响到蒸腾作用的强弱,是影响瞬时液流的最主要因子,由冬季至夏季,光照时间逐渐延长,光照强度逐渐增强,从而使得树干液流维持在高值水平的时间逐渐增加,由夏季至冬季则反之。
由于树干液流的大小决定于生物学结构、土壤供水状况和气象因子,同一树种不同个体之间以及不同树种之间的边材和心材面积比率差异很大, 即使具有相同或相似胸径, 其液流速率差异也很大(孙鹏森等,2000)。冬、春、夏、秋季杉木日液流量分别为(0.32±0.26),(3.22±2.94),(7.31±3.95)和(2.18±1.10) kg·d-1,远小于尖峰栲(树高17.6 m,DBH为38.6 cm,边材面积587.6 cm2)雨季(103.5 kg·d-1)和旱季(41.3 kg·d-1)的单株平均液流量(张刚华等,2007),生长季杉木日液流量是华北落叶松(熊伟等,2003)的2倍以上。
回归分析表明,树干液流与环境因子的关系随研究时间尺度的不同而不同,影响瞬时液流速率的环境因子主要是辐射和气温,影响日液流量的主要环境因子是气温、土壤水分含量和辐射,影响月液流量的主要环境因子为冠上气温和相对湿度。在本研究地域,冬季和春季的阴雨天气较多且气温较低,土壤表层含水量较丰富,液流耗水主要由表土供应;夏季和秋季的晴朗天气多且气温高,表层土壤含水量不足以维持杉木的蒸腾耗水,蒸腾耗水主要由深层土壤供给。
在本研究中,由于只有一套TDP探针,只能对杉木标准木的液流特征作初步的分析研究,在今后的研究中,需增加TDP探针的数目,更全面地研究杉木液流的空间变化,并通过尺度转换研究杉木林分蒸腾特征。
白云岗, 宋郁东, 周宏飞, 等. 2007. 热脉冲法对胡杨树干液流的监测与蒸腾过程的模拟. 水土保持学报, 21(3): 188-192. DOI:10.3321/j.issn:1009-2242.2007.03.042 |
常学向, 赵文智. 2004. 荒漠绿洲农田防护树种二白杨生长季节树干液流的变化. 生态学报, 24(7): 1436-1441. DOI:10.3321/j.issn:1000-0933.2004.07.018 |
刘健, 赖娜娜, 赵炳祥, 等. 2007. 银杏树体茎流变化及其对环境因子的响应. 中国农学通报, 23(6): 232-237. DOI:10.3969/j.issn.1000-6850.2007.06.052 |
刘奉觉, 郑世锴, 巨关升, 等. 1997. 树木蒸腾耗水测算技术的比较研究. 林业科学, 33(2): 117-126. DOI:10.3321/j.issn:1001-7488.1997.02.003 |
马玲, 赵平, 饶兴权, 等. 2005. 马占相思树干液流特征及其与环境因子的关系. 生态学报, 25(9): 2145-2151. DOI:10.3321/j.issn:1000-0933.2005.09.003 |
马李一, 孙鹏森, 马履一. 2001. 油松、刺槐单木与林分水平耗水量的尺度转换. 北京林业大学学报, 23(4): 1-5. |
聂立水, 李吉跃, 翟洪波. 2005. 油松、栓皮栎树干液流速率的比较. 生态学报, 25(8): 1934-1940. DOI:10.3321/j.issn:1000-0933.2005.08.015 |
司建华, 冯超, 张小由, 等. 2007. 极端干旱区荒漠河岸林胡杨生长季树干液流变化. 中国沙漠, 27(3): 442-447. DOI:10.3321/j.issn:1000-694X.2007.03.016 |
孙慧珍, 周晓峰, 赵惠勋. 2002. 白桦树干液流的动态研究. 生态学报, 22(9): 1387-1391. DOI:10.3321/j.issn:1000-0933.2002.09.003 |
孙慧珍, 孙龙, 王传宽, 等. 2005. 东北东部山区主要树种树干液流研究. 林业科学, 41(3): 36-42. DOI:10.3321/j.issn:1001-7488.2005.03.006 |
孙鹏森, 马履一, 王小平, 等. 2000. 油松树干液流的时空变异性研究. 北京林业大学学报, 22(5): 1-6. DOI:10.3321/j.issn:1000-1522.2000.05.001 |
孙守家, 古润泽, 丛日晨, 等. 2006. 银杏树干茎流变化及其对抑制蒸腾措施的响应. 林业科学, 42(5): 22-28. |
王华田, 赵文飞, 马履一. 2006. 侧柏树干边材液流的空间变化规律及其相关因子. 林业科学, 42(7): 21-27. |
肖文发, 徐德应, 刘世荣, 等. 2002. 杉木人工林针叶光合与蒸腾作用的时空特征. 林业科学, 38(5): 38-46. DOI:10.3321/j.issn:1001-7488.2002.05.007 |
谢恒星, 张振华, 杨润亚, 等. 2007. 龙爪槐树干液流相对于气象因子的滞后效应分析. 林业科学, 43(5): 106-110. |
熊伟, 王彦辉, 徐德应. 2003. 宁南山区华北落叶松人工林蒸腾耗水规律及其对环境因子的响应. 林业科学, 39(2): 1-7. |
严昌荣, DowneyA, 韩兴国, 等. 1999. 北京山区落叶阔叶林中核桃楸在生长中期的树干液流研究. 生态学报, 19(6): 793-797. DOI:10.3321/j.issn:1000-0933.1999.06.005 |
尹光彩, 周国逸, 王旭, 等. 2003. 应用热脉冲系统对桉树人工林树液流通量的研究. 生态学报, 23(10): 1984-1990. DOI:10.3321/j.issn:1000-0933.2003.10.005 |
张刚华, 陈步锋, 聂洁珠, 等. 2007. 热带山地雨林尖峰栲边材液流及其与环境因子的关系. 应用生态学报, 18(4): 742-748. DOI:10.3321/j.issn:1001-9332.2007.04.006 |
张小由, 康尔泗, 司建华, 等. 2006. 胡杨蒸腾耗水的单木测定与林分转换研究. 林业科学, 42(7): 28-32. |
赵平, 饶兴权, 马玲, 等. 2006. 基于树干液流测定值进行尺度扩展的马占相思林段蒸腾和冠层气孔导度. 植物生态学报, 30(4): 655-665. DOI:10.3321/j.issn:1005-264X.2006.04.016 |
赵文飞, 王迎, 王华田, 等. 2007. 不同季节麻栎树干贮水量的动态变化. 林业科学, 43(4): 115-120. |
Baker J M, Van Bavel H M. 1987. Measurement of mass flow of water in the stems of herbaceous plants. Plant, Cell and Environment, 10: 777-782. |
Bovard B D, Curtis P S, Vogel C S, et al. 2005. Environmental controls on sap flow in a northern hardwood forest. Tree Physiology, 25: 31-38. DOI:10.1093/treephys/25.1.31 |
Gazal R M, Scott R L, Goodrich D C, et al. 2006. Controls on transpiration in a semiarid riparian cottonwood forest. Agricultural and Forest Meteorology, 137: 56-67. DOI:10.1016/j.agrformet.2006.03.002 |
Granier A. 1985. A new method of sap flow measurement in tree stems. Annals of Forest Science, 42(2): 193-200. DOI:10.1051/forest:19850204 |
Granier A. 1987. Sap flow measurement in Douglas-fir tree trunks by means of a new thermal method. Annals of Forest Science, 44(1): 1-14. DOI:10.1051/forest:19870101 |