 2009, Vol. 45
2009, Vol. 45文章信息
- 霍常富, 孙海龙, 王政权, 范志强, 赵晓敏.
- Huo Changfu, Sun Hailong, Wang Zhengquan, Fan Zhiqiang, Zhao Xiaomin
- 光照和氮营养对水曲柳苗木生长及碳-氮代谢的影响
- Effects of Light and Nitrogen on Growth, Carbon and Nitrogen Metabolism of Fraxinus mandshurica Seedlings
- 林业科学, 2009, 45(7): 38-44.
- Scientia Silvae Sinicae, 2009, 45(7): 38-44.
-
文章历史
- 收稿日期:2007-12-03
-
作者相关文章



光是绿色植物叶片进行光合作用的唯一能量来源,氮(N)是植物根系吸收最多的矿质元素之一,两者是影响植物生长发育和演化的最重要环境因子(关义新等,2000).森林中光和N的分布很不均匀,生长于不同光照强度(光强)和N有效性环境中的植物,其叶片碳-氮代谢过程将发生很大程度的改变(Makino et al., 1997).研究表明,在高光强环境中,植株叶面积减小,叶片变厚,光合产物和可溶性蛋白含量较丰富,N同化关键酶(硝酸还原酶,NR)活性较高(Crabtree et al., 1993;Vos et al., 2000;关义新等,2000).在弱光环境中生长的植株通常向茎和叶分配较多的碳(生物量),提高冠根比,以便获取更多的光能,因此,单株叶面积和叶质量比较大,但叶片中光合产物积累减少,NR活性也降低,硝酸盐在叶片中积累(Crabtree et al., 1993; Makino et al., 1997;Vos et al., 2001;关义新等,2000).N对植株生长也有明显影响.N供给充足的植株生长较快,叶片可溶性蛋白含量较高,氮代谢活动旺盛,NR活性较高,而硝酸盐含量较低(Pinto et al., 1999;吴楚等,2003).N缺乏通常使植株的生长受到抑制,生理代谢活动减慢(de Pinheiro et al., 2000).高等植物叶片把光合碳同化和N同化集于一体,二者之间既存在激烈的竞争又有着十分紧密的联系.因此,只有叶片内碳代谢和氮代谢相互协调,才能使植株处于最佳的生长状态(侯光炯等,1982).
在温带森林生态系统中,光照和氮营养由于季节和林内微环境的变化,它们在时间和空间上都存在很大的异质性(Chapin Ⅲ et al., 2002;韩有志等,2004),而且二者经常共同影响植物的生长发育(Crabtree et al., 1993;霍常富等,2008a).水曲柳(Fraxinus mandshurica)是我国东北地区主要的造林树种之一,在北方温带森林生态系统中具有十分重要的地位.因其材质优良、价格昂贵,过度的开发和利用使其森林面积锐减.水曲柳甚至被列为濒危种(王义弘等,1994),急需对其保护和恢复.以往研究发现,水曲柳林下更新幼苗在林隙处明显多于林内,而且林隙处1年生幼苗较多,但2~3年生幼苗和幼树数量急剧减少(臧润国,1999;韩有志等,2004),这是否与林隙处光强和N有效性增加有关还不清楚.因此,本文针对光强和N浓度2种环境条件,通过分析水曲柳苗木生物量积累、叶片生长以及叶片碳-氮代谢指标的变化,来探讨水曲柳苗木对不同光、N环境条件的生理适应机制,旨在为水曲柳林分更新以及优良苗木的培育提供理论依据.
1 材料与方法 1.1 植物材料及培养试验采用温室砂培方法,在东北林业大学林木遗传育种基地(哈尔滨实验林场内)进行.苗木为来自吉林省红石林业局培育的同一批1年生水曲柳幼苗,从中随机选出均匀一致的苗木192株,于2006年5月1日栽植到塑料桶(直径30 cm,高28 cm)中,每桶4株.培养基质为河砂,先将河砂用清水浸泡,洗去泥土,再经0.5%盐酸浸泡24 h,然后用自来水冲洗至中性.栽植后保证苗木充足的水分,待苗木全部成活后供给Hoagland完全营养液,具体配法参照吴楚等(2003)的方法.每3天浇1次营养液(每次每桶200 mL),营养液浇灌时间为早上8:00—9:00,同时保证苗木充足的水分,浇水时间为早上8:00—9:00和下午4:00—5:00分2次进行.温室内昼夜平均温度分别为30 ℃和20 ℃,相对湿度85%以上,光照平均14 h·d-1.
1.2 试验设计于6月10日,进行光强和N浓度处理.试验设2种光强处理:全光照(温室内自然光照)和遮荫(温室内自然光照的40%).遮荫处理采用黑色遮荫网在苗木正上方1 m处进行遮荫,使网下光强为室内自然光强的40%左右.每种光强处理下再设置4个供N水平:1,4,8和16 mmol·L-1(分别记为N1,N4,N8和N16).N浓度处理,只改变营养液中NH4NO3的浓度,其他营养成分保持不变.本试验共8个处理,每个处理6桶(即6次重复),共计48桶.
1.3 植株收获及指标测定处理3个月后(9月5日)进行全株收获.将苗木从桶中小心取出,洗净根系,迅速拿回实验室进行指标测定和化学分析.用枝剪把苗木根和茎分开,摘下叶片(保持复叶),称量根、茎和叶鲜质量.叶面积由激光叶面积仪(CI-203,美国)测定.取一部分鲜叶用于可溶性蛋白、硝酸盐含量和硝酸还原酶活性的测定,剩余样品置于80 ℃烘箱中烘72 h,取出称量.计算冠根比(地上部分干质量/根干质量,Crown mass/Root mass,C/R),比叶面积(总叶面积/植株总干质量,Specific leaf area,SLA),叶质量比(叶干质量/植株总干质量,Leaf mass ratio, LWR).
1.4 化学分析可溶性蛋白含量测定,采用考马斯亮蓝G-250染色法(李合生,2000).硝酸盐含量测定,采用水杨酸-硫酸比色法(李合生,2000).可溶性糖和淀粉含量测定,根据Buysse等(1993)的方法.称取40 mg干样(粉末)在10 mL 80% (v/v)的乙醇中过夜提取,离心(5 000 r·min-1,10 min),将上清液倒入50 mL容量瓶中,定容,用于可溶性糖分析.将残余物烘干,用5 mL 3%(w/w)的盐酸煮沸3 h,过滤转移到100 mL容量瓶中,用于淀粉含量分析.分析过程为,吸取1 mL提取液,加入1 mL 28%(w/w)的苯酚(溶于80%乙醇中),然后立即加入5 mL浓硫酸,迅速摇晃1 min,静止15 min后,于490 nm处比色.标准曲线,由葡萄糖(60 ℃,烘24 h)溶液配制.
硝酸还原酶(NR)活性的测定,根据Knoepp等(1993)的方法.称取0.5 g切碎的叶片,放入盛有20 mL缓冲溶液(含40 mmol·L-1 NO3-、0.05 mol·L-1 Tris和1.5%丙醇,pH值7.5)的烧杯中.在切碎的叶片上覆盖塑料网,以保证叶片组织完全浸入溶液中.将烧杯放在干燥器中,通气3次,每次1~2 min,然后将烧杯放在水浴(25 ℃)中,暗孵育1 h.孵育之后,吸取2份1 mL样品溶液,加入1 mL 1%(w/v)磺胺和1 mL 0.02%(w/v)二盐酸萘乙二胺(这2种物质均溶于1.5 mol·L-1盐酸中),用分光光度计在540 nm处测定NO2-的含量.
1.5 数据分析把光强(2水平)和N浓度(4水平)看作2因素,以生物量、叶面积和可溶性蛋白等指标为因变量,由SPSS 11.5 for windows软件进行双因素方差分析,并进行光、N及其交互作用的显著性检验(α= 0.05).叶片碳-氮代谢各指标之间相关性分析也由SPSS完成,表中数值为Pearson相关系数,在α= 0.05水平上进行双尾检验.多重比较采用邓肯法(Duncan’s multiple range test).图表中数据为6个重复的平均值±标准误差,图由Sigmaplot 9.0制作完成.
2 结果与分析 2.1 光强和氮浓度对水曲柳苗木生物量和叶片生长的影响与全光照相比,遮荫处理使苗木比叶面积、冠根比、单株叶面积和叶质量比分别增加了35.5%,24.9%,21.8%和18.5%,但是苗木单株生物量下降了19.6%(表 1).除苗木比叶面积外,N供给浓度对其余生长参数都产生明显影响(表 1).无论在全光照还是遮荫条件下,苗木单株生物量和叶面积均随N供给浓度的增加而显著提高(P<0.05).而冠根比和叶质量比,只有在遮荫条件下随N供给浓度的增加而显著提高(P<0.05),但在全光照条件下,差异未达显著水平.
|
|
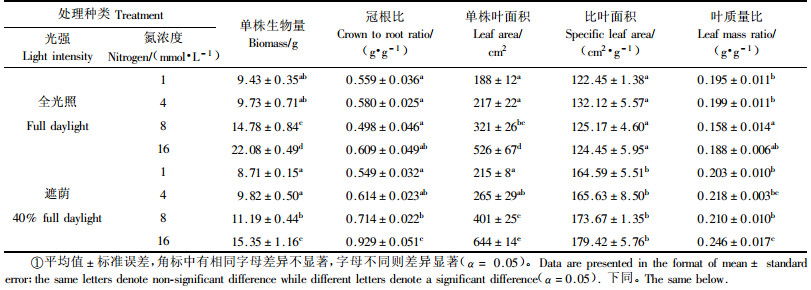
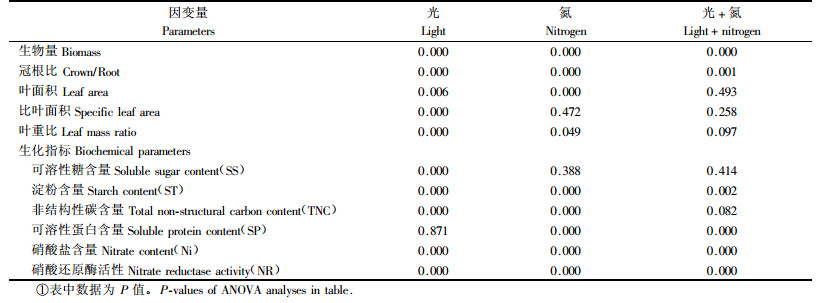
方差分析表明,光强处理对苗木各生长参数影响均达极显著水平(P<0.01),N处理除对比叶面积无明显影响以外,对其余指标的影响也都达到显著水平(P<0.05).光和N对苗木单株生物量和冠根比有明显的交互作用(表 2).
|
|
遮荫处理显著降低了叶片中可溶性糖、淀粉和非结构性碳水化合物(TNC)含量(图 1),随供N浓度的增加叶片中淀粉和TNC含量显著降低(P<0.05)(图 1B, C).不考虑光照条件,低N时(N1),淀粉在叶片内大量积累,平均值达到389 mg·g-1,在N4时其含量急剧下降至282 mg·g-1,N8和N16时略有回升(图 1B).由于水曲柳苗木叶片中淀粉含量是可溶性糖含量的近3倍,因此,TNC含量的变化趋势与淀粉类似(图 1C).此外,光和氮对苗木叶片淀粉含量有极显著的交互作用(P<0.01,表 2).

|
图 1 光强和氮浓度处理对水曲柳苗木叶片可溶性糖含量、淀粉含量和非结构性碳水化合物含量的影响 Figure 1 Effects of light intensity and nitrogen concentration on soluble sugar content, starch content and TNC content in leaves of Ash tree seedlings |
光强和氮浓度及其交互作用对苗木叶片3个氮代谢指标都产生了显著影响(P<0.05)(图 2,表 2).遮荫使叶片硝酸盐含量和硝酸还原酶(NR)活性显著降低(P<0.05,图 2B, C),但使叶片可溶性蛋白含量显著提高(P<0.05,图 2A).随N供给水平的提高,可溶性蛋白含量和NR活性明显上升(图 2A, C),硝酸盐含量则显著下降(图 2B).值得指出的是,在遮荫条件下,N供给充足(N16)时,叶片可溶性蛋白含量略有下降(图 2A),同时硝酸盐在叶片中积累(图 2B).此外,在全光照条件下,NR活性随N供给水平的提高增加较快(回归直线斜率k=57.6),而在遮荫时则较慢(回归直线斜率k=13.5)(图 2C).

|
图 2 光强和氮浓度处理对水曲柳苗木叶片可溶性蛋白含量、硝酸盐含量和硝酸还原酶活性的影响 Figure 2 Effects of light intensity and nitrogen concentration on soluble protein content, nitrate content and NR activity in leaves of Ash tree seedlings |
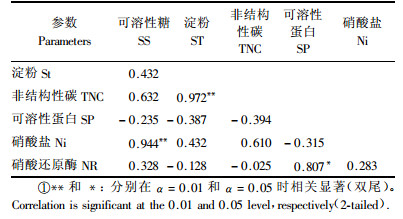
相关分析表明(表 3),在叶片碳代谢指标中,TNC含量与淀粉含量之间的相关系数最大(r=0.972,P<0.01),这也是叶片中TNC与淀粉含量变化趋势(图 1B, C)相一致的主要原因.在叶片氮代谢指标中,可溶性蛋白(SP)与NR活性显著相关(r=0.807, P<0.05),而硝酸盐(Nitrate,Ni)与SP和NR都不具有显著相关性,这可能是由于叶片中Ni含量不仅取决于叶片对其还原能力,还取决于根系对其吸收能力.水曲柳苗木叶片碳-氮代谢指标之间,只有叶片中可溶性糖(SS)含量与Ni含量具有显著的正相关性(r=0.944,P<0.01),其他指标之间相关性未达显著水平(表 3).总的来看,叶片中SP和NR活性分别与碳代谢指标(SS,ST和TNC)之间具有一定程度的负相关性(表 3).
|
|
植物最大的特点是生根后便不能自由移动,只能通过调节其形态和生理过程来适应外界多变的环境条件,这是植物长期适应环境的策略(臧润国,1999;Chapin Ⅲ et al., 2002).当光照不足时,植物通常将光合作用固定的C优先分配于地上部分,使冠根比、叶质量比、总叶面积和比叶面积增加,这些地上部分形态特征的改变,有利于其获得更多的光能,适应光照不足的环境(Sims et al., 1994;Vos et al., 2001).本研究发现,遮荫处理使苗木冠根比、叶质量比、总叶面积和比叶面积极显著增加(P<0.01,表 1、2),表明1年生水曲柳幼苗对光照不足的环境具有较强的调节和适应能力.虽然植物对低光照条件具有一定的适应能力,但是光照不足仍会导致叶片光合速率下降,光合产物积累减少,并最终影响植物总生物量(Vos et al., 2001;霍常富等,2008a).在本研究中,遮荫处理使水曲柳幼苗单株生物量平均下降了19.6%(P<0.01,表 1,表 2),这表明水曲柳生长对光的依赖性较强.在许多自然生态系统中,N常常是植物生长发育的限制因子(Crawford et al., 1998).植株体内N的获取依赖于C的投入,当植株在N缺乏时,根系生物量相对较大以便获取更多的N来维持其生长,因此冠根比较小;而在N充足时,为了获取更多的光能,地上部分生长迅速,使冠根比增大(Masarovicova et al., 2000).本研究发现,水曲柳苗木冠根比在全光照条件下受N供给浓度的影响较小,而在遮荫条件下随N供给浓度的提高而显著增大(P<0.05),这可能是因为遮荫条件下光照是水曲柳生长的主要限制因子,随着N供给浓度的增大,这种限制也越大,所以导致植株向地上部分分配更多的生物量,以获得更多的光.虽然水曲柳苗木冠根比在全光照下受N供给影响较小,但是其单株生物量和叶面积随供N浓度的增大而显著增加(P<0.05),这表明水曲柳是对N比较敏感的树种(吴楚等,2003).
可溶性糖是叶片碳运输和代谢过程中主要形式,淀粉是叶片内碳积累的主要形式(武维华,2003),它们是衡量植株体内碳代谢的重要指标.植株体内可溶性糖和淀粉含量受外界环境条件(光照、N浓度和水分等)的影响较大(Kathju et al., 2001;Makino et al., 1997;张莉等,2003).本文研究结果表明,遮荫处理使水曲柳苗木叶片可溶性糖和淀粉含量显著下降(P<0.05),这可能是遮荫使苗木叶片光合速率下降的直接结果(霍常富等,2008b).然而,在N严重缺乏时,叶片可溶性糖和淀粉含量反而升高(图 1),这与其他植株在遭受盐胁迫和水分胁迫时的研究结果相似(Balibrea et al., 2000;张莉等,2003).造成这种现象的原因可能有2个,一方面与植株在逆境条件下的特殊生理反应有关(张莉等,2003),另一方面是由于植株在逆境条件下生长相对缓慢,对光合产物的需求相应减少,从而使大量的可溶性糖和淀粉在叶片中积累(Munns,1993),后者可能是造成本试验中水曲柳苗木叶片可溶性糖和淀粉积累的主要原因,其生物量在N缺乏下显著降低从另一个侧面也说明了这一点.
植物体内氮代谢主要包括无机N的还原、同化及有机N化合物转化和合成等过程(Miller et al., 2004).光和N对水曲柳苗木N代谢都有显著影响(图 2),遮荫使叶片可溶性蛋白含量(在N16时)和NR活性明显下降,表明水曲柳苗木对N的还原和同化受到了抑制.这主要是因为N的还原和同化需要消耗大量的能量,而苗木处于遮荫条件下所提供的能量非常有限,最终导致叶片中合成的可溶性蛋白减少.然而,值得指出的是,在遮荫条件下并没有发现硝酸盐在水曲柳苗木叶片中大量积累,这可能是遮荫使苗木根系吸收硝酸盐的能力下降的结果,但还需要进一步通过试验来证实.N是植物体内蛋白质、叶绿素和酶类等许多物质的重要组分(武维华,2003),因此N供给浓度的高低对植物体内可溶性蛋白含量和NR活性具有显著影响(Kathju et al., 2001;Miller et al., 2004).本研究得出,随N供给浓度的增加苗木叶片可溶性蛋白含量和NR活性显著提高(P<0.05),有利于苗木对N的还原和同化,因此叶片中硝酸盐积累较少.然而,在遮荫条件下高N(N16)处理时硝酸盐含量略有回升,这可能与遮荫条件下叶片NR活性上升相对缓慢有关.NR是一种诱导酶,而诱导过程需要光的积极参与(Miller et al., 2004),遮荫使叶片NR活性明显降低,而且随N供给浓度的增加上升的速度也较全光照条件下慢,这表明水曲柳苗木叶片NR活性受光强影响很大,进而影响蛋白质合成乃至整个氮代谢过程.因此,可以说光强和N浓度处理对叶片NR活性的影响是叶片N代谢变化的关键.
碳-氮代谢是植物体内最基本的两大代谢过程,碳代谢为N代谢提供能量和碳骨架,氮代谢同碳代谢竞争光反应生成的同化力(ATP和NADPH)及其代谢中间产物,因此碳-氮代谢在植物体内具有重要的生理关系(武维华,2003).二者之间既存在着激烈的竞争,又有着十分紧密的联系,因此植物如何协调植物体内碳-氮代谢的关系对其生长发育具有重要影响(侯光炯等,1982;田纪春等,2001).本研究结果表明,水曲柳苗木叶片碳-氮代谢具有一定的负相关性(表 3),即叶片中可溶性糖、淀粉和TNC含量随可溶性蛋白和NR活性的增加而减少,这与关义新等(2000)对玉米(Zea mays)叶片代谢关系的研究结果相一致.此外,叶片可溶性糖含量与硝酸盐含量具有极显著的正相关性,而叶片中较高的硝酸盐含量意味着其还原同化过程可能受阻,因此,可以说叶片中维持较高的可溶性糖含量是以牺牲叶片N同化为代价的结果.
综上所述,1年生水曲柳苗木对光强和N营养的改变反应都十分敏感.虽然水曲柳幼苗对弱光和低N环境具有一定的适应和调节能力,但是光照不足和N贫乏最终还是使苗木的生长发育受到严重抑制.因此,在我国东北森林的经营和管理过程中,要特别注重营造适合水曲柳幼苗生长的林下光强和土壤有效性N等环境.
关义新, 林葆, 凌碧莹. 2000. 光、氮及其互作对玉米幼苗叶片光合和碳、氮同化的影响. 作物学报, 26(6): 806-812. DOI:10.3321/j.issn:0496-3490.2000.06.026 |
韩有志, 王政权, 谷加存. 2004. 林分光照空间异质性对水曲柳更新的影响. 植物生态学报, 28(4): 468-475. DOI:10.3321/j.issn:1005-264X.2004.04.004 |
霍常富, 王政权, 孙海龙, 等. 2008a. 光照和氮交互作用对水曲柳苗木生长、生物量和氮分配的影响. 应用生态学报, 19(8): 1658-1664. |
霍常富, 孙海龙, 王政权, 等. 2008b. 光照和氮营养对水曲柳苗木光合特性的影响. 生态学杂志, 27(8): 1255-1261. |
侯光炯, 周源芳. 1982. 农业土壤学基础知识. 成都: 四川人民出版社, 81-84.
|
李合生. 2000. 植物生理生化实验原理和技术. 北京: 高等教育出版社, 123-124, 184-185.
|
田纪春, 王学臣, 刘广田. 2001. 植物的光合作用与光合氮、碳代谢的耦联及调节. 生命科学, 13(4): 145-147. DOI:10.3969/j.issn.1004-0374.2001.04.001 |
王义弘, 柴一新, 慕长龙. 1994. 水曲柳的生态学研究. 东北林业大学学报, 22(1): 1-6. |
吴楚, 王政权, 范志强, 等. 2003. 不同氮浓度和形态比例对水曲柳幼苗叶绿素合成、叶片总氮含量、光合作用以及生物量分配的影响. 植物生态学报, 27(6): 771-779. DOI:10.3321/j.issn:1005-264X.2003.06.007 |
武维华. 2003. 植物生理学. 北京: 科学出版社, 91-92.
|
张莉, 续九如. 2003. 水分胁迫下刺槐不同无性系生理生化反应的研究. 林业科学, 39(4): 162-167. DOI:10.3321/j.issn:1001-7488.2003.04.028 |
臧润国. 1999. 林隙动态与森林生物多样性. 北京: 中国林业出版社, 30-32.
|
Balibrea M E, Amico J D, Bolarín M C, et al. 2000. Carbon partitioning and sucrose metabolism in tomato plants growing under salinity. Physiologia Plantarum, 110: 503-511. DOI:10.1111/ppl.2000.110.issue-4 |
Buysse J, Merckx R. 1993. An improved colormetric method to quantify sugar content of plant tissue. Journal of Experimental Botany, 44: 1627-1629. DOI:10.1093/jxb/44.10.1627 |
Chapin Ⅲ F S, Matson P A, Mooney H A. 2002. Principles of terrestrial ecosystem ecology. New York: Spinger-Verlag, 197-223.
|
Crabtree R C, Bazzaz F A. 1993. Seedling response of four birch species to simulated nitrogen deposition: ammonium vs. nitrate. Ecological Applications, 3(2): 315-321. DOI:10.2307/1941834 |
Crawford N M, Glass A D M. 1998. Molecular and physiological aspects of nitrate up take in plants. Trends in Plant Science, 3: 381-395. |
de Pinheiro H A R, Marcells L F M. 2000. Regulation of growth at steady-state nitrogen nutrition in Lettuce (Lactuca sativa L.): interactive effects of nitrogen and irradiance. Annals of Botany, 86: 1073-1080. DOI:10.1006/anbo.2000.1268 |
Kathju S, Burman U, Garg B K. 2001. Influence of nitrogen fertilization on water relations, photosynthesis, carbohydrate and nitrogen metabolism of diverse pearl millet genotypes under arid conditions. J Agricultural Science, Cambridge, 137: 307-308. |
Knoepp J D, Turner D P, Tingey D T. 1993. Effects of ammonium and nitrate on nutrient uptake and activity of nitrogen assimilating enzymes in western hemlock. Forest Ecology and Management, 59: 179-191. DOI:10.1016/0378-1127(93)90001-4 |
Makino A, Sato T, Nakano H, et al. 1997. Leaf photosynthesis, plant growth and nitrogen allocation in rice under different irradiances. Planta, 203: 390-398. DOI:10.1007/s004250050205 |
Masarovicova E, Welschen R, Lux A, et al. 2000. Photosynthesis, biomass partitioning and peroxisomicine A1 production of Karwinskia species in response to nitrogen supply. Physiologia Plantarum, 108: 300-306. DOI:10.1034/j.1399-3054.2000.108003300.x |
Miller A J, Cramer M D. 2004. Root nitrogen acquisition and assimilation. Plant and Soil, 274: 1-36. |
Munns R. 1993. Physiological processes limiting plant growth in saline soils: Some dogmas and hypotheses. Plant Cell and Environment, 16: 15-24. DOI:10.1111/pce.1993.16.issue-1 |
Nakaji T, Fukami M, Dokiya Y. 2001. Effects of high nitrogen load on growth, photo synthesis and nutrient status of Cryptomeria japonica and Pinus densiflora seedlings. Trees, 15: 453-461. |
Pinto M E, Casati P, Hsu T P, et al. 1999. Effects of UV-B radiation on growth, photosynthesis, UV-B-absorbing compounds and NADP-malic enzyme in bean (Phaseolus vulgaris L.)grown under different nitrogen conditions. Journal of Photochemistry Photobiology B: Biology, 48: 200-209. DOI:10.1016/S1011-1344(99)00031-7 |
Sims D A, Gebauer R L E, Pearcy R W. 1994. Scaling sun and shade photosynthetic acclimation of Alocasia macrorrhiza to whole-plant performance:Ⅱ. Simulation of carbon balance and growth at different photon flux densities. Plant Cell and Environment, 17: 889-900. DOI:10.1111/pce.1994.17.issue-8 |
Vos J, van der Putten P E L. 2001. Effects of partial shading of the potato plant on photosynthesis of treated leaves, leaf area expansion and allocation of nitrogen and dry matter in component plant parts. European Journal of Agronomy, 14: 209-220. DOI:10.1016/S1161-0301(00)00090-3 |