 2009, Vol. 45
2009, Vol. 45文章信息
- 王鹏程, 肖文发, 姚婧, 张守攻, 黄志霖, 曾立雄, 潘磊.
- Wang Pengcheng, Xiao Wenfa, Yao Jing, Zhang Shougong, Huang Zhilin, Zeng Lixiong, Pan Lei
- 三峡库区3种典型森林主要组成树种的种群结构及更新
- Population Structure and Regeneration of the Main Tree Species in Three Typical Forests in the Three Gorges Reservoir Area
- 林业科学, 2009, 45(7): 7-15.
- Scientia Silvae Sinicae, 2009, 45(7): 7-15.
-
文章历史
- 收稿日期:2007-12-24
-
作者相关文章


2. 中国林业科学研究院森林生态环境与保护研究所 北京 100091;
3. 湖北省林业科学研究院 武汉 430079
2. Institute of Forest Ecology, Environment and Protection, CAF Beijing 100091;
3. Forestry Science Institute of Hubei Province Wuhan 430079
植被动态是植被生态学研究的热点问题之一,而群落演替是植被动态研究的核心内容.近年来,植被动态研究领域不断扩大,内涵也扩展到任何尺度上的时空变化(周小勇等,2005;丁圣彦等,2004).目前研究群落动态的方法主要有建立永久固定样地进行定位研究、以空间代替时间进行比较研究及通过数学模型进行动态模拟研究等,由于以空间代替时间进行比较研究方法不受研究时间的限制而广泛应用(Johnson, 1997;Rebertus et al., 1993;刘峰等,2000).种群是群落的组成部分,其结构能客观地体现群落的发展趋势.因此,研究群落种群结构对阐明种群生态特性、更新对策乃至群落的形成及其稳定性与演替规律等都具有重要意义(Wu et al., 2002; 达良俊, 2004).优势种群在群落中占据主导地位,其种群结构动态可以反映群落结构变化.通过不同演替阶段群落优势种群结构动态分析确定树种更新类型,可以推测群落演替趋势,而种群年龄结构是描述种群数量动态的重要指标,由于在野外调查过程中,森林群落个体年龄难以准确测定,常采用大小级结构代替年龄结构研究种群动态(Johnson, 1997; Rebertus et al., 1993;彭少麟等,1998).
三峡库区虽经过多年的林业生态工程建设,但大部分退化群落仍处于自然恢复状态,普遍存在演替更新困难问题,制约了区域生态建设和社会经济的可持续发展(肖文发等,2000).本研究以三峡库区典型森林类型为研究对象,以空间代替时间,从森林主要优势种群的年龄结构入手,通过对库区内3种典型森林类型主要组成树种的种群结构及其更新机制的研究,诊断群落所处的演替阶段及其发展方向,为三峡库区退化植被恢复与地带性植物群落重建提供理论依据.
1 研究区概况三峡库区地处106°14′—111°28′ E、28°56′—31°44′ N,属中亚热带湿润地区.由于受季风的影响,冬季雨水少而夏季雨水集中,年降水量1 000~1 200 mm,高于同纬度的中国东部地区,年平均气温为15~18 ℃,土壤以红、黄壤为主,同时还有较大面积的紫色土、石灰土.山脉多呈西北向东南倾斜状,长江由西向东呈30°~60°倾角切断了巫山和大巴山余脉,强烈的水蚀切割作用形成特殊岩溶地貌类型.三峡库区自然环境优越,具有我国东西与南北两大生物界过渡的特点,亚热带至寒温带植被极为丰富,主要包含寒温性针叶林、温性针叶林、暖温性针叶林、阔叶林、竹林、灌丛和灌草丛及灌草丛等7种植被型(肖文发等,2000).
2 研究方法 2.1 样地设置与调查在研究区内布设代表性强的针叶林、阔叶林、针阔混交林样地各10块,样地面积为20 m×20 m,并在样地内梅花状布设5个2 m×2 m的小样方.调查样地乔木种、树高和胸径及其幼树幼苗数量;同时调查小样方灌木层和草本层的物种、高度、株数和盖度.调查时间为2004年7—8月和2005年4—5月,调查样地基本信息见表 1.
|
|

优势种是群落的重要组成部分,创造特定的群落环境并影响其他物种的组成及分布,因而优势种的确定对研究群落特征有着重要的意义.本文通过计算和比较样地乔木层组成树种重要值,从而确立群落优势种、伴生种.乔木层物种重要值的计算公式为:物种重要值=(相对密度+相对频度+相对显著度)×100/3.
2.3 群落演替与更新分析利用每木检尺调查数据,采取胸径级结构代替年龄结构,绘制径级频率图分析种群径级频率分布,以此判断种群动态趋势.种群的径级频率图中纵坐标表示样方内某一物种的个体数,横坐标表示样方内该物种的胸径径阶,其中径阶是采取5刻度为单位,使用上限排外法分为(0,5),[5,10),[10,15),[15,20),[20,25)和≥25等6个刻度范围(刘峰等,2000;丁圣彦,1999),各树种可归纳为5个种群结构类型,即单峰型(unimodal type)、间歇型(sporadic type)、逆J字型(inverse-J type)、L型(L type)和单柱型(unibar type)(丁圣彦等,2003;达良俊等,2004),在这里“峰”指在某一径阶范围内呈现的、具有中心顶点的、抛物线型的胸径级频率分布形状.
各种群结构类型主要特征及对应的更新类型如下:1)单峰型:只有少数大胸径级个体出现,径级频率分布呈现一至数个不连续的单峰状径级,但缺乏小径阶的后继更新个体,种群的发展趋势属衰退类型;2)间歇型:群落内个体数较多,但中径级个体数较少,径级频率分布呈不连续状,有多个单峰出现,同时具有较多的小径级后继更新个体,种群的发展趋势属间歇性发展型;3)逆J字型:个体的径级分布为连续分布,并呈现出2个峰,即大径级处的小峰和小径级处的大峰,小径级后继个体较多,种群的发展趋势属成熟稳定类型;4) L型:径级频率呈连续分布,个体集中于小径阶处,无大径级个体,种群的发展趋势属成长类型;5)单柱型:只有在小径级处有个体分布和出现,无明显的增长趋势.
3 结果与分析 3.1 3种森林类型典型样地乔木层物种组成及重要值针叶林样地乔木层优势种主要包括巴山松(Pinus henryi)、华山松(Pinus armandi)、马尾松(Pinus massoniana)、杉木(Cunninghamia lanceolata)、柏木(Cupressus funebris)等针叶树种,其重要值为67.37~100.00;伴生树种主要为樟树(Cinnamomum camphora)、化香(Platycarya strobilacea)、麻栎(Quercus acutissima)、栓皮栎(Quercus variabilis)、四照花(Cornus kousa var.chinensis)等阔叶树种,少量样地分布有柏木、杉木等针叶树种,重要值为1.47~24.35,见表 2.
|
|
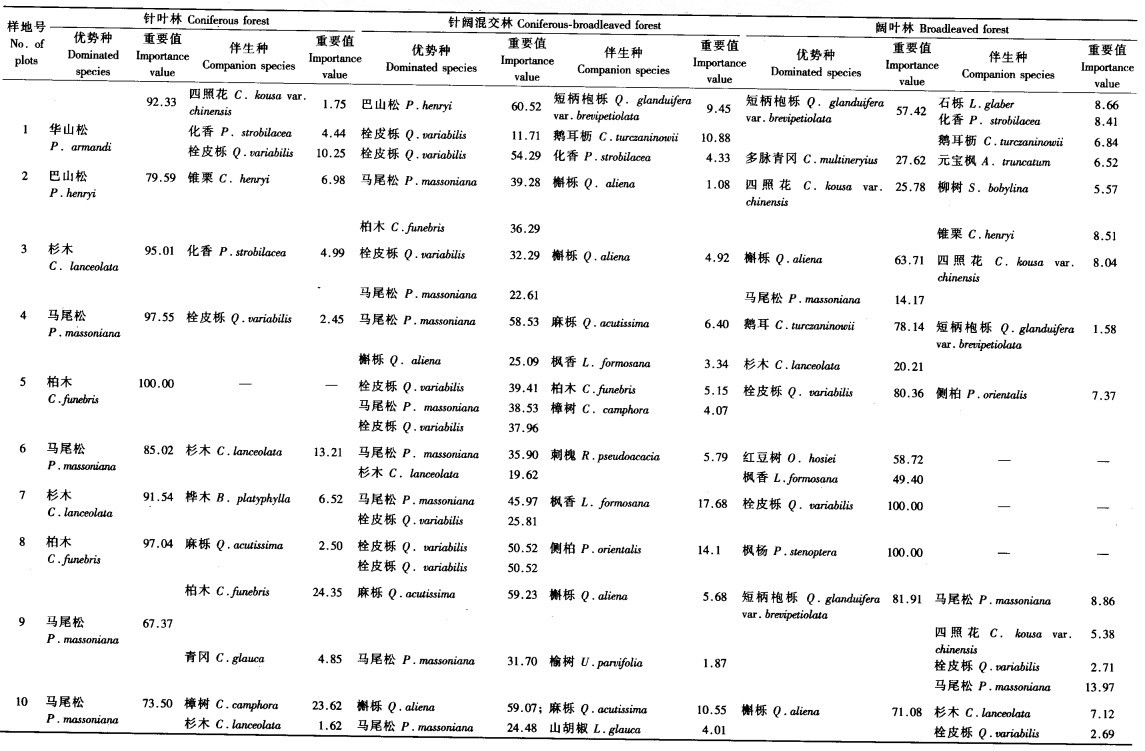
由表 2可以看出,针阔混交林样地乔木层主要树种包括巴山松、马尾松、柏木、杉木、侧柏(Platycladus orientalis)等针叶树种和栓皮栎、短柄NFDA6栎(Quercus glanduifera var. brevipetiolata)、化香、槲栎(Quercus aliena)、麻栎、鹅耳栎(Carpinus turczaninowii)等阔叶树种.针阔混交林样地的优势树种同时包含1个以上针叶树种和阔叶树种,其单一树种的重要值在11.71~60.52,伴生树种多以枫香(Liquidambar formosana)等阔叶树种为主,重要值均在17.68以下.
在10个阔叶林样地中,各样地包含1个或多个阔叶树种优势种,树种主要为短柄NFDA6栎、多脉青冈(Cyclobalanopsis multineryius)、四照花、槲栎、栓皮栎、红豆树(Ormosia hosiei)、枫香、枫杨(Pterocaya stenoptera)等,树种的重要值为25.78~100.00.伴生树种主要为四照花、化香、鹅耳栎、青冈(Cyclobalanopsis glauca)、锥栗(Castanea henryi)等阔叶树种和杉木、马尾松、侧柏等针叶树种,见表 2.
3.2 森林垂直结构三峡库区森林在垂直方向上可分为乔木层、灌木层和草本层.针叶林乔木层物种数较少,以马尾松、杉木、柏木、巴山松和华山松为主,林分郁闭度为0.50~0.80,其中松林郁闭度为0.60~0.80,高于柏木(0.40~0.75)和杉木林(0.50~0.70).针叶林乔木树种个体高度级频度顶点为10~15 m,伴生种多处于亚乔木层,以壳斗科植物为主,个体高度级频度顶点处于5~10 m间.在3种森林类型中,阔叶林树种数量最多,主要为壳斗科植物,林分郁闭度为0.60~0.90,个体高度级频度顶点出现在5~10 m.针阔混交林乔木层个体高度级频度顶点在5~10 m之间.
灌木层物种较丰富,但不同样地种类组成差别较大,毛黄栌(Cotinus coggygria var. pubescens)、檵木(Loropetalum chinense)、盐肤木(Rhus chinensis)、野蔷薇(Rosa multiflora)分布较广.灌木层盖度一般低于0.20,高度为0.5~3.5 m之间.草本层植物种类较多,且各样地物种基本相同,主要物种为菝葜(Smilax china)、苔草(Fagus lucicla)、泽兰(Eupatorium japonicum)、芒(Miscanthus sinensis)、蕨类(Pteridophyta)等,盖度一般在0.10以下.
3.3 不同类型森林的优势种年龄结构及更新模式 3.3.1 阔叶林优势种年龄结构及更新模式库区阔叶林可分为优势种不明显群落(2号和6号样地)和优势种明显群落(其他阔叶群落样地)2类,见表 3.优势种明显群落的优势种以短柄NFDA6栎、槲栎、栓皮栎等壳斗科植物为主,伴生种的科属比较繁多,主要为化香、鹅耳栎、四照花等.
|
|

从表 3看出,在短柄NFDA6栎为优势种的群落中,优势种在各个径级均有较多个体存在,可以预测,在环境稳定的情况下,短柄NFDA6栎种群结构将比较稳定,在群落演替的后期也将会处于优势地位.从种群结构类型来划分,短柄NFDA6栎种群个体数多,且小径级范围内的后续更新个体数量也比较多,表明该种群为稳定型,其胸径频率分布属于逆J字型(1,9号样地).以槲栎(3,10号样地)、栓皮栎(5,7号样地)为优势种的群落中槲栎、栓皮栎的胸径频率分布属于逆J字型,种群为稳定发展类型.群落其他树种,如石栎、鹅耳栎、化香等只能作为伴生树种在群落中存在,化香胸径频率分布属于L型,种群处于发展状态;石栎为逆J字型;而马尾松和锥栗属于单峰衰退类型,在群落演替末期可能被其他树种取代.

|
图 1 乔木层主要物种高度级频度分布 Figure 1 The height class frequency distribution of main species in the arbor layer |
在优势种不明显的群落中(2,6号样地),树种较多且分布比较均匀,优势种为多脉青冈、四照花、红豆树等(表 3).四照花大径级所占比例较多,小径级范围内株数较少,其种群结构偏向于衰退型,其胸径频率分布表现为小径级株树较少的单峰型特征.多脉青冈属于成熟型,胸径分布频率为逆J字型;尾叶山茶(Camellia caudata)和交让木(Daphniphyllum macropodum)则是多数个体处于小径级范围内,其中交让木属于典型“L型”增长类型,尾叶山茶种群则属于逆J字型的发展类型.
其他阔叶林群落如鹅耳栎群落(4号样地)、枫杨群落(8号样地)中鹅耳栎和枫杨的胸径频率分布分别属于间歇型、逆J字型,鹅耳栎种群为间歇性发展型,枫杨种群为成熟稳定类型.在鹅耳栎群落中,作为伴生种的杉木种群呈L型,有一定发展趋势,但杉木喜阳,由于鹅耳栎占据乔木层上层,导致杉木大径个体存活数量有限,无法占据主导地位.
从阔叶林优势种的径级结构可以看出,虽然各群落物种组成不同,但阔叶群落中的优势种种群结构保持稳定,短期内不会发生优势种的更新取代.
3.3.2 针阔混交林优势种年龄结构及更新模式从表 4看出,三峡库区针阔混交林中针叶树种低海拔地区以马尾松为主,高海拔地区则以巴山松为主,其他伴生针叶树种为柏木、侧柏等;阔叶树种主要为短柄NFDA6栎、槲栎、麻栎、栓皮栎、化香、四照花等.
|
|
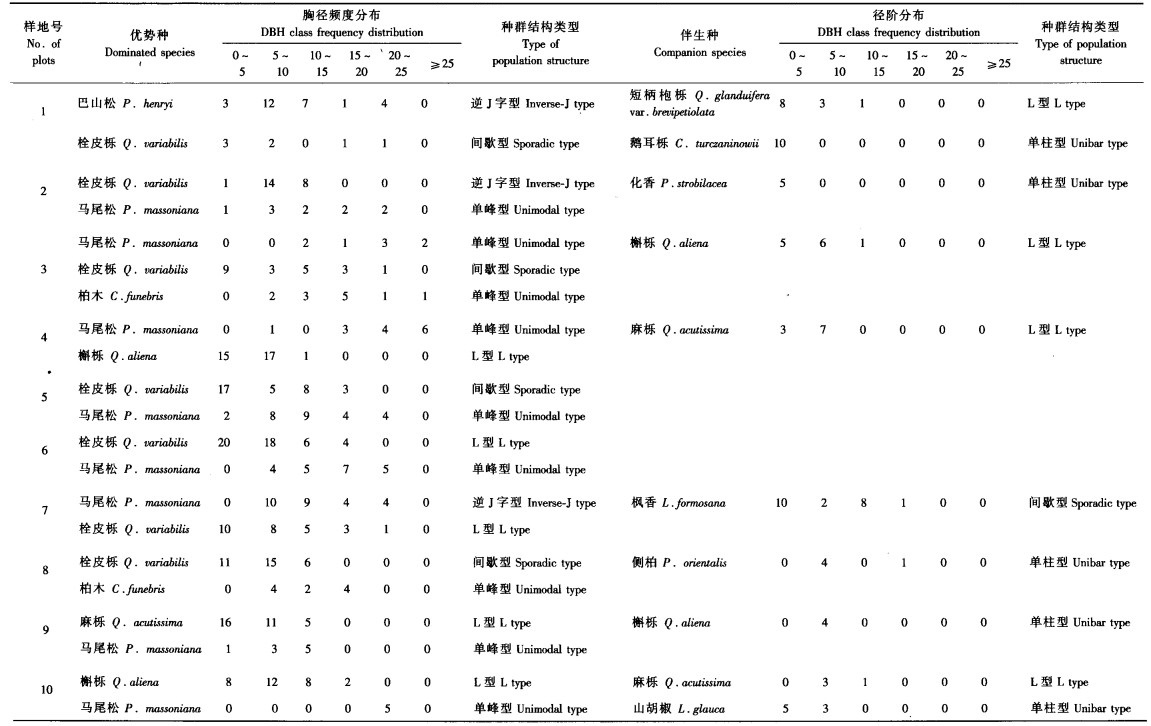
巴山松-短柄NFDA6栎群落(1号样地)中优势种巴山松胸径频率属于逆J字型,种群结构将保持稳定;短柄NFDA6栎属于逐渐增长类型,胸径频率分布符合L型特征;而栓皮栎属于间歇型,胸径频度分布为多峰状态.在马尾松与壳斗科植物为优势种的混交群落中,除7号样地外,其他样地的马尾松种群处于衰退的单峰型,基本无小径阶个体,只残留较小比例的大径级个体.柏木种群结构短期内变动不大,其胸径分布属于单峰型.针阔混交林中栓皮栎的小径阶个体数很多,但中间径级的个体存在小峰值,可预测栓皮栎的发展具不连续性,为偏向于增长的间歇型或L型.槲栎和麻栎合计个体数占乔木层总株数80%,且个体大部分处于具很大发展趋势的(0,10)径级内,胸径频率分布属稳定增长的L型.作为阔叶伴生种的鹅耳栎、化香等都是明显的单柱型,树种只在小径阶范围内存在零星个体,在大径阶部分个体基本灭绝,其种群发展趋势为不明显的增长趋势.
总体来看,针阔混交林中大部分针叶树种优势种的更新趋势为衰退型,如柏木、马尾松;阔叶优势树种都为种群结构发展稳定的逆J字型或间歇发展型,如槲栎、栓皮栎、短柄NFDA6栎等.
3.3.3 针叶林主要优势种年龄结构及更新模式三峡库区针叶林优势种以马尾松、杉木、柏木为主,在高海拔地区则以华山松、巴山松为主,而伴生种多是以化香、四照花、栓皮栎、麻栎、樟树等阔叶树种为主,不同样地伴生种有所不同,见表 5.
|
|

从表 5可以看出,杉木林、柏木林、马尾松林的针叶树种年龄结构单一,个体都集中在大径级范围内,几乎无小径级后续更新个体,属于典型的单峰型,因此种群在更新后期会逐渐衰退.华山松林(1号样地)灌木层中出现较多化香、山胡椒和四照花等阔叶乔木幼树,优势种华山松的径级结构类型属于单峰型,在小径级的后续个体极少,种群呈现衰退趋势;化香和山胡椒种群均为不明显性增长的单柱型,而四照花种群为逐渐增长的L型.巴山松林(2号样地)优势种地位十分明显,巴山松种群在大径级范围内个体数很多,而在小径级处极少,仅在灌木层偶见少量幼树,为成熟型种群的特点,胸径频率分布属于后续更新减少的多峰型,伴生种的胸径频率分布为L型,种群呈增长趋势.
总体来看,库区针叶林的优势种种群结构均以衰退型或以间歇性发展类型为主,针叶树种更新较弱,如杉木和华山松等为衰退单峰型树种.增长型树种都为阔叶树种,如化香、四照花和栓皮栎等树种的种群结构为增长的L型.
4 结论与讨论三峡库区针叶林中优势种为强阳性树种,种群的个体集中分布在大径级中,占据了群落的上层,随着林分郁闭度升高,林内光照减弱造成更新受阻,只有少量个体残留于超高层,研究结论与苏志尧等(2000)的研究结果一致,因此,强阳性树种的更新需要强度干扰和较大面积裸地的出现;阳性阔叶树种子通过林窗入侵针叶林,萌发后迅速占据林冠层,在演替的末期均取代针叶树种(苏志尧等, 2000).落叶阔叶树种的生活习性都为喜光的阳性树种,在演替过程过程中会出现针阔混交群落阶段.可以初步预测三峡库区针叶林类林的演替趋势为针叶林→以针叶树种为主的针阔混交群落→以阳性阔叶树种为主的针阔混交林.
在马尾松和柏木等先锋树种为主的针阔混交林演替早期,强阳性针叶树种占据林冠层,为耐荫树种提供适宜的萌发和生长环境,而自身种子萌发和幼苗生长受到限制,种群为单峰的衰退型.巴山松为优势树种的针阔混交林中巴山松为人工栽培,强烈和频繁的人工干扰影响林下物种的入侵和生长,而亚优势种或伴生种如短柄NFDA6栎、化香、栓皮栎、槲栎、麻栎等阳性落叶阔叶树种都为增长的L型和稳定的逆J字型.这种结构特征正好验证了针叶林演替过程.虽然不同样地乔木层的针叶树种和阔叶树种的径级结构不完全相同,但具有共同特征,即针叶树种种群为衰退型种群,阔叶树种总存在一个或多个增长型种群,表明不同样地处于不同演替阶段.
在阔叶林的林冠层和超高层有少数残留阳性针叶先锋树种,阳性阔叶树种的幼树幼苗具有一定的耐荫性,在林荫下进行天然更新,完成种群自身构筑,并逐渐占据主林层(苏志尧等, 2000).不同样地树种的生态学特性和群落环境决定了组成树种的种群发展趋势,针叶树种种群处于衰退的末期,多数阳性阔叶树种在前期生长迅速,随着林分郁闭度增大,阳性阔叶树种生长和更新受阻,中性的阔叶树种个体数目逐渐增多,最终形成以中性树种为主的常绿阔叶林(Tang et al., 2002).
在三峡库区,保存着部分以马尾松、杉木和日本落叶松等针叶树种为主的用材林以及荒山造林形成的大面积针叶林,森林健康状况受到普遍重视,因此,在未来经营管理中应围绕抚育阔叶树种这一目标进行调控,在低海拔地区实施轮封,加强幼树幼苗的保护;在高海拔地区,选择和保护优良的阔叶树母树,扩大阔叶树种入侵通道——林窗的面积和密度,保证足够数量的阔叶树种幼苗,促进种群发展,以快速完成针叶林向顶极群落演替,增强森林生态系统的稳定性(张文辉等,2008).
通过种群结构类型推断树种的更新类型和群落的变化趋势已成为研究森林群落演替的重要手段,但在研究中采取径级的个体数代替年龄结构,忽略群落不同个体因局部生境条件不同而导致的生长差异,这种误差会影响演替趋势的预测精度.
丁圣彦, 宋永昌. 1999. 浙江天童常绿阔叶林演替序列优势种光合生理生态的比较. 生态学报, 19(3): 318-323. DOI:10.3321/j.issn:1000-0933.1999.03.005 |
丁圣彦, 宋永昌. 2004. 常绿阔叶林植被动态研究进展. 生态学报, 24(8): 1769-1779. |
丁圣彦, 宋永昌. 2003. 演替研究在常绿阔叶林抚育和恢复上的应用. 应用生态学报, 14(3): 423-426. DOI:10.3321/j.issn:1001-9332.2003.03.022 |
丁圣彦, 宋永昌. 1998. 常绿阔叶林演替过程中马尾松消退的原因. 植物学报, 40(8): 755-760. DOI:10.3321/j.issn:1672-9072.1998.08.011 |
达良俊, 杨永川, 宋永昌. 2004. 浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型. 植物生态学报, 28(3): 376-384. DOI:10.3321/j.issn:1005-264X.2004.03.013 |
刘峰, 陈伟烈, 贺金生. 2000. 神农架地区锐齿槲栎种群结构与更新的研究. 植物生态学报, 24(4): 396-401. DOI:10.3321/j.issn:1005-264X.2000.04.003 |
彭少麟, 方炜, 任海. 1998. 鼎湖山厚壳桂群落演替过程的组成和结构动态. 植物生态学报, 22(3): 245-249. DOI:10.3321/j.issn:1005-264X.1998.03.007 |
桑卫国, 陈灵芝, 于顺利, 等. 2000. 蒙古栎红松林物种组成和结构动态的研究. 植物生态学报, 24(2): 231-237. DOI:10.3321/j.issn:1005-264X.2000.02.018 |
苏志尧, 吴大荣, 陈北光. 2000. 粤北天然林优势种群结构与空间格局动态. 应用生态学报, 11(3): 337-341. DOI:10.3321/j.issn:1001-9332.2000.03.004 |
王永健, 陶建平, 李媛, 等. 2007. 华西箭竹对卧龙亚高山森林不同演替阶段物种多样性与乔木更新的影响. 林业科学, 43(2): 1-7. |
肖文发, 李建文, 程瑞梅, 等. 2000. 长江三峡库区陆生动植物生态. 重庆: 西南师范大学出版社.
|
张文辉, 卢彦昌, 周建云, 等. 2008. 巴山北坡不同干扰条件下栓皮栎种群结构与动态. 林业科学, 44(7): 11-16. DOI:10.3321/j.issn:1001-7488.2008.07.003 |
周小勇, 黄忠良, 欧阳学军, 等. 2005. 鼎湖山季风常绿阔叶林原锥栗-厚壳桂-荷木群落演替. 生态学报, 25(1): 37-44. DOI:10.3321/j.issn:1000-0933.2005.01.007 |
Johnson J B. 1997. Stand structure and vegetation dynamics of a subalpine treed fen in Rocky Mountain National Park, Colorado. Journal of Vegetation Science, 8: 337-342. DOI:10.2307/3237322 |
Rebertus A J, Veblen T T. 1993. Structure and tree-fall gap dynamics of old-growth Nothofagus forests in Tierra del Fuego, Argentina. Journal of Vegetation Science, 4: 641-654. DOI:10.2307/3236129 |
Tang C Q, Ohsawa M. 2002. Coexistence mechanisms of evergreen, deciduous and coniferous trees in a mid-montane mixed forest on Mt. Emei, Sichuan, China. Plant Ecology, 161: 215-230. DOI:10.1023/A:1020395830795 |
Wu X P, Zheng Y, Ma K P. 2002. Population distribution and dynamics of Quercus liaotungensis, Fraxinus rhynchophlla and Acer mono in Dongling Mountain, Beijing. Acta Botanica Sinica, 44(2): 212-223. |