 2007, Vol. 43
2007, Vol. 43文章信息
- 王玉魁, 闫艳霞, 慈龙骏, 王保平, 崔令军, 杨超伟.
- Wang Yukui, Yan Yanxia, Ci Longjun, Wang Baoping, Cui Lingjun, Yang Chaowei.
- 7种滨藜的解剖结构及C4光合特征
- Anatomical Structure and C4 Photosynthetic Characteristics of Seven Atriplex Species
- 林业科学, 2007, 43(增刊1): 72-76.
- Scientia Silvae Sinicae, 2007, 43(增刊1): 72-76.
-
文章历史
- 收稿日期:2007-04-30
-
作者相关文章



2. 中国林业科学研究院林业研究所 北京 100091;
3. 北京林业大学水土保持学院水土保持与荒漠化防治重点实验室 北京 100083
2. Research Institute of Forestry, CAF Beijing 100091;
3. College of Soil and Water Conservation, Beijing Forestry University Beijing 100083
滨藜属(Atriplex)植物是干旱、半干旱地区的典型植物, 具有耐旱、耐寒、抗盐碱等特性, 是盐碱地改良和荒漠化土地治理的优良树种之一(李梅, 2002)。滨藜属包括180多个草本和灌木植物种, 广泛分布于温带和亚热带地区, 以美国西北部种类最多(刘英, 2002)。1965年Kortschack等以甘蔗为试材观察到CO2固定的初期产物是四碳酸, Hatch等(1966)在甘蔗研究上获得证实并提出了C4途径。随着植物解剖技术提高以及稳定碳同位素技术提高的应用, 大量C4植物被人们发现(韩家懋等, 2002; 蒋高明等, 2004)。有关C4植物生理、生化特性近几年才有所阐明(Hans et al., 2005)。滨藜属是藜科(Chenopodiaceae) C4植物分布较集中的属(韩家懋等, 2002; 蒋高明等, 2004), 但目前对滨藜属植物光合作用途径的系统研究未见报道。
不同光合途径的植物具有不同的δ13C值(Farquhar et al., 1989; 韩家懋等, 2002; 苏培玺等, 2005)。Bender(1968; 1971)最早提出用稳定碳同位素技术区分植物的光合途径, 确定C3植物的δ13C值为-23 ‰~ -32 ‰, C4植物为-6 ‰~ -19 ‰, 其他学者的研究结果也大都在这个范围(Farquhar et al., 1982; Hattersley, 1982; Farquhar, 1983)。潘瑞炽等(2001)和蒋高明等(2004)认为, 典型C4植物在叶的解剖结构中具有较大的维管束鞘薄壁细胞, 其中含有叶绿体; 维管束鞘的外侧密接一层环状或近于环状排列的叶肉细胞, 组成“花环型”(Kranz type)结构。但有研究表明, 并非所有的C4植物都具有花环结构, 如Voznesenskaya等(2001)发现异子蓬(Borszczowia aralocaspica)有C4植物光合特征, 但没有花环结构。Smith等(1973)认为植物叶片δ13C可以作为花环结构的指示, δ13C值在-9 ‰~ -16 ‰之间植物具有花环结构, 在-23 ‰~ -32 ‰之间无花环结构。本文通过对7种滨藜属植物的叶解剖结构观察、稳定碳同位素测定和光合特征综合分析, 旨在对其光合作用途径进行判别。
1 材料与方法 1.1 试验区概况甘肃省民勤治沙试验站地处巴丹吉林沙漠与腾格里沙漠的交接地带(102° 58′ E, 38° 34′ N, 海拔1 375 m), 属温带大陆性荒漠气候区。年平均气温7.6 ℃, 极端低温-30.8 ℃, 极端高温40.0 ℃, 年均降水量113.2 mm, 年均蒸发量2 604.3 mm, 年日照时数2 799.4 h, 相对湿度47 %, 干燥度4.5以上, 无霜期175 d, 年平均大风沙日数83 d, 多集中在3 —5月, 年平均风速2.5 m·s-1。地带性土壤为灰棕荒漠土和风沙土, pH 8.5左右, 地下水位20 m, 矿化度0.51 ~ 1.5 g·L-1。
国家林业局泡桐研究开发中心实验基地(113° 34′—113° 52′ E, 34° 53′—35° 05′ N)在河南原阳县境内, 属暖温带大陆性季风气候区。年平均气温14.4 ℃, 1月平均气温-0.6 ℃; 年降水量582.9 mm, 其中60 %集中在6 —8月份, 年蒸发量为2 034 mm; 年平均日照时数2 324.5 h; 无霜期224 d。土壤为黄河冲积风沙土, pH 7.5 ~ 8.5, 地下水位10 ~ 12 m。
1.2 试验材料试验材料为2003年从北美洲引进的7种滨藜: Atriplex canescens、A.confertifolia、A.corrugata、A. gardneri、A.lentiformis、A.gardneri trident和A.obovata, 对照植物有:沙棘(Hippophae rhamnoides)、刺槐(Robinia pseudoacacia)、新疆杨(Populus alba var.pyramidalis)、梭梭(Haloxylon ammodendron)、沙拐枣(Calligonum mongolicum)、玉米(Zea mays)和同期从新疆引种到民勤的白滨藜(A. cana)。
1.3 试验方法 1.3.1 叶片解剖方法取3年生滨藜当年生成熟叶片(阳面顶部第3叶片), 用FAA固定液固定(50 %酒精: 5 %福尔马林: 5 %冰醋酸=90: 5: 5) (V/V)。试材经系列酒精脱水后, 常规石蜡切片法制片, 用旋转式切片机(Rotary Microtome, Leitz, Germany)切片(厚度8 μm), 番红-固绿对染, 二甲苯透明, 阿拉伯树胶封片(李正理等, 1981), 在显微镜下观察拍照。
1.3.2 δ13C和Δ值测定2006年植物生长季节, 分别在民勤和原阳试验区采样。自植株顶部向下取第1 ~ 10片成熟叶子, 每种植物各选1 ~ 3株混合制样, 70 ℃烘48 h, 粉碎过筛。由核工业部三所质谱分析室使用Delta plus XP同位素比例质谱仪测定稳定性碳同位素比率(δ13C), 仪器分析精确度为0.03 ‰。每种植物供试样品重复制备测定2次, 以国际标准物质PDB(Pee Dee Belemnite)为标准, 用下列公式计算δ13C值(Kloeppel et al., 1998):

|
(1) |
式中: Rsample是植物叶片样品的13C/12C比率; Rstandard是测定过程中标准物质PDB的13C/12C比率。
空气样用1 000 mL真空玻璃细劲瓶采集, CO2通过真空分离线提纯后在质谱仪上测定。碳同位素辨别力(Δ)用下列公式计算(Kloeppel et al., 1998):
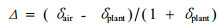
|
(2) |
每个种各选择3株有代表性的滨藜为标准木, 树冠顶端向下第3 ~ 5片充分展开的叶片为被测样本。用Li -6400光合分析仪在0 ~ 2 500 μmol·m-2s-1光合有效辐射(PAR)范围内, 利用人工光源(LED)设定0、10、50、100、200、500、1 000、1 500、2 000、2 500 μmol·m-2s-1等强度, 测定各叶片的净光合速率(Pn)。测试时间选在植物生长季(8 —9月)的晴天进行, 气温(35.21 ±2.35)℃, 大气CO2浓度(394.85 ± 12.48)μmol·mol-1, 参比室空气湿度控制在50 %±6.52 %。
1.4 数据处理数据处理和分析采用Excel 2005和SPSS11.5软件。滨藜叶片Pn对PAR的响应关系采用(Bassman et al., 1991)经验方程进行拟合:
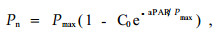
|
(3) |
式中, Pmax为最大净光合速率(μmol CO2·m-2s-1), a和C0为弱光下净光合速率趋近于零的常数, PAR为光合有效辐射, Pn为特定PAR条件下的净光合速率。
通过适合性检验, 若方程良好(p < 0.05), 分别计算光补偿点LCP =Pmax ln(C0)/a、光饱和点LSP = Pmaxln(100 ×C0)a和表观量子效率AQY=aC0。
2 结果与分析 2.1 叶的解剖结构特征7种滨藜叶的横切面基本结构相同, 都是由叶肉细胞、上下表皮、栅栏组织、维管束及维管束鞘等组成。叶表皮细胞外侧均有数层排列疏松的叶肉细胞, 这些细胞没有角质层, 易脱落, 具有保护、储水和减少蒸腾作用。上下表皮细胞横切面为扁平状, 外侧有角质层, 内侧为排列紧密、柱状、单层的栅栏组织细胞, 气孔在上下表皮均有分布。叶片横切面的主脉上有4个内始式维管束, 其外围有1层半封闭的维管束鞘细胞(bundle sheath cell), 也称花环细胞(Kranz cell), 叶片内小的维管束仅由导管和筛管组成, 其外被大且排列紧密的维管束鞘细胞, 包围成花环状, 细胞内含有大量的叶绿体(图版Ⅰ-1 ~ 8)。

|
图版Ⅰ Plate Ⅰ |
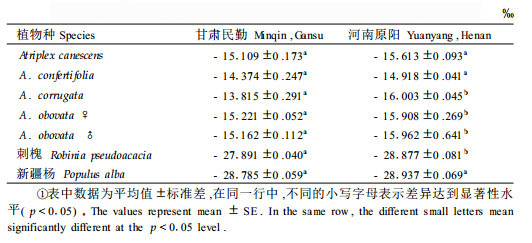
由表 1可知, 测试的8种滨藜属植物除白滨藜外, 其他7种滨藜的稳定碳同位素比率比较接近, δ13C值在-13.815 ‰~ -15.613 ‰之间, 属C4植物的δ13C值范围内(Bender, 1968; 1971;Smith et al., 1971; 1973);所测得的14种植物叶片或同化枝的δ13C值之间存在极显著差异(p < 0.01), 多重比较(Duncan法)显示: A. corrugata与C4植物梭梭和沙拐枣(苏培玺, 2005)的δ13C值比较接近, 差异不显著(p < 0.05);6种滨藜A.lentiformis、A. confertifolia、A. gardneri、A. obovata、A.gardneri trident和A. canescens的δ13C值种间差异不显著; 白滨藜与C3植物刺槐和新疆杨的δ13C值接近, 差异不显著(p >0.05)。以稳定性碳同位素辨别力(Δ)对各种植物的差异显著性检验结果与δ13C值相同, 7种滨藜属植物的Δ值在3.514 ‰~ 5.346 ‰之间, 与C4植物玉米、梭梭和沙拐枣的Δ值相近, 均小于6 ‰, 白滨藜与C3植物沙棘、刺槐和新疆杨的Δ值比较接近, 均大于16 ‰。δ13C值越大, Δ值越小, 反之亦然。
|
|

表 2显示, 在植物生长季节, 同期引种的滨藜δ13C值在民勤干旱沙区和高于相对湿润的原阳; C3植物刺槐和新疆杨的δ13C值在原阳同样低于在民勤。这一趋势与韩家懋等(2002)的研究相符, 即不同环境无论C3植物还是C4植物, 它们的δ13C值由东往西逐渐偏正。多重比较结果表明, 新疆杨、A.canescens、A. confertifolia的δ13C值地点间差异不显著(p >0.05);刺槐、A. corrugata、A. obovata ♀及A. obovata ♂地点间差异显著(p < 0.05)。
|
|
在原阳试区测定了7种滨藜的净光合速率对光强的响应, 以刺槐为对照, 计算得到的光合生理参数见表 3。结果显示, 7种滨藜的光饱和点均在1 603 ~ 2 185 μmol·m-2s-1之间, C3植物刺槐为1 357 μmol·m-2s-1。C4植物的光饱和点高, 而C3植物的光饱和点较低, Larcher(1995)报道, C4植物的光饱和点大于1 500 μmol·m-2 s-1, C3植物在1 000 ~ 1 500 μmol·m-2s-1之间, 本文研究结果与Larcher的结果一致。苏培玺等(2005)研究认为C4植物的表观量子效率在0.04 mol CO2·mol-1 photons以上, 本试验测试的7种滨藜的表观量子效率在0.049 5 ~ 0.071 8 mol CO2·mol-1 photons之间, 属于C4植物的范围, 且均显著高于C3植物刺槐(0.027 4 mol CO2· mol-1 photons)。
|
|
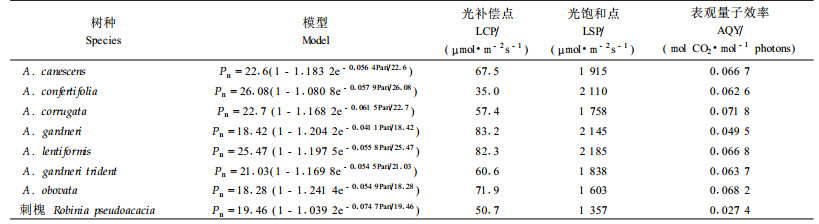
研究结果表明, 7种滨藜属植物叶片的横切面均由外层叶肉细胞和内层维管束鞘细胞构成明显的花环结构, 维管束鞘细胞内含叶绿体; 植物生长季节分别在干旱沙区(民勤)和亚湿润区(原阳)测定的δ13C值在-13 ‰~ -16 ‰之间; 光饱和点均大于1 600 μmol·m-2s-1, 表观量子效率在0.04 μmol·mol-1以上。证明引进的7种滨藜均为C4植物, 其光合作用途径没有随地理位置的变化而改变。
7种滨藜的光补偿点在35.0 ~ 83.2 μmol·m-2s-1之间; 苏培玺等(2005)的研究结果表明C3植物柠条具有较高的光补偿点(137 μmol·m-2·s-1), 显著高于C4植物, 但本文测试的C3植物刺槐光补偿点较低(50.7 μmol·m-2 s-1), 所以作者认为光补偿点能否作为判别植物光合作用途径的指标还需进一步研究。
新生代晚期出现的C4植物是地球环境演化的产物(韩家懋等, 2002), 是植物长期进化对环境的适应。由于环境条件对植物光合作用途径的影响(牛书丽等, 2004), 使鉴别复杂化。植物叶片δ13C值易受叶片气体交换调控的生理机制的影响, 光照、大气压力、温度、湿度等均有可能引起δ13C值的变化(陈拓, 2003)。所以, 对植物光合作用途径的鉴别应尽量采用多种手段, 结合环境条件分析进行鉴别。随着人类对光合作用途径研究的深入, 对于不同光合作用途径的鉴别手段日渐完善, 目前主要鉴别方法有:形态解剖、稳定碳同位素、酶学研究以及14CO2示踪。
陈拓, 杨梅学, 冯虎元, 等. 2003. 青藏高原北部植物叶片碳同位素组成的空间特征. 冰川冻土, 25(1): 83-87. DOI:10.3969/j.issn.1000-0240.2003.01.013 |
韩家懋, 王国安, 刘东生. 2002. C4植物的出现与全球环境变化. 地球前缘, 9(1): 233-243. |
Hans L, Chapin F S, Pons T L.2005.植物生理生态学.张国平, 周伟军, 译.杭州: 浙江大学出版社, 36-44
|
蒋高明, 常杰, 高玉葆, 等. 2004. 植物生理生态学. 北京: 高等教育出版社, 59-64.
|
李梅. 2002. 征服沙尘暴的神奇植物——美国滨藜. 农业环境与发展, (4): 19. |
李正理, 李荣敖. 1981. 我国甘肃九种旱生植物同化枝的解剖观察. 植物学报, 23(3): 181-185. |
刘英. 2002. 抗旱耐盐四翅滨藜. 植物杂志, (4): 16-17. |
牛书丽, 蒋高明, 李永庚. 2004. C3与C4植物的环境调控. 生态学报, 24(2): 308-314. |
潘瑞炽, 王小菁, 李娘辉. 2001. 植物生理学. 4版. 北京: 高等教育出版社, 80-85.
|
苏培玺, 安黎哲, 马瑞君, 等. 2005. 荒漠植物梭梭和沙拐枣的花环结构及C4光合特征. 植物生态学报, 29(1): 1-7. DOI:10.3321/j.issn:1005-264X.2005.01.001 |
孙双峰, 黄建辉, 林光辉, 等. 2005. 稳定同位素技术在植物水分利用研究中的应用. 生态学报, 25(9): 2362-2371. DOI:10.3321/j.issn:1000-0933.2005.09.032 |
Bassman J B, Zwier J C. 1991. Gas exchange characteristics of Populus trichocarpa, Populus deltoides and Populus trichocarpa ×P. deltoides clone.Tree Physiol, 8: 145-149. DOI:10.1093/treephys/8.2.145 |
Bender M M. 1968. Mass spectrometric studies of carbon-13 variation in corn and other grasses. Radiocarbon, 10: 468-472. DOI:10.1017/S0033822200011103 |
Bender M M. 1971. Variations in the 13C/12C rations of plants in relation to the pathway of photosynthetic carbon dioxide fixation. Phytochemistry, 10: 1239-1244. DOI:10.1016/S0031-9422(00)84324-1 |
Farquhar G D, O'Leary M H, Berry J A. 1982. On the relationship between carbon isotope discrimination and intercellular carbon dioxide concentration in leaves. Australian Journal of Plant Physiology, 9: 121-137. |
Farquhar G D. 1983. On the nature of carbon isotope discrimination in C4 species. Australian Journal of Plant Physiology, 10: 205-226. |
Farquhar G D, Ehleringer J R, Hubick K T. 1989. Carbon isotope discrimination in photosynthesis. Annual Review of Plant Physiology and Molecular Biology, 40: 503-537. DOI:10.1146/annurev.pp.40.060189.002443 |
Hatch M D, Slack C R. 1966. Photosynthesis by sugar cane leaves-a new carboxylation reaction and the pathway of sugar formation. Biochem J, 101: 103-111. DOI:10.1042/bj1010103 |
Hattersley P W. 1982. δ13C values of C4 types in grasses. Australian Journal of Plant Physiology, 9: 139-154. |
Kloeppel B D, Gower S T, Treichel I W, et al. 1998. Foliar carbon isotope discrimination in Larix species and sympatric evergreen conifers:a global comparison. Oecologia, 114: 153-159. DOI:10.1007/s004420050431 |
Kortschack H P, Hartt E E, Burr G O. 1965. Carbon dioxide fixation in sugarcare leaves. Plant Physiology, 40: 209-213. DOI:10.1104/pp.40.2.209 |
Larcher W. 1995. Physiological plant ecology:ecopysiology and stress physiology of functional groups. 3rd ed. New York: Springer, Berlin, Heidelberg,, 94-111.
|
Nier A O, Gulbransen E A. 1939. Variations in the relative abundance of the carbon isotopes. Journal of the American Chemical Society, 61: 697-698. DOI:10.1021/ja01872a047 |
Park R, Epstein S. 1960. Carbon isotope fractionation during photosynthesis. Geochimica Et. Cosmochimica Acta, 21: 110-126. DOI:10.1016/S0016-7037(60)80006-3 |
Park R, Epstein S. 1961. Metabolic fractionation of 13C and 12C in plants. Plant Physiology, 36: 133-138. DOI:10.1104/pp.36.2.133 |
Sage R F, Li M R, Monson R K. 1999.The taxonomic distribution of C4 Photosynthesis∥Sage R F, Russell K M. C4 Plant Biology. San Diego: Academy Press, 551-584
|
Smith B N, Epstein S. 1971. Two categories of 13C/12C rations for higher plants. Plant Physiology, 47: 380-384. DOI:10.1104/pp.47.3.380 |
Smith B N, Brown W V. 1973. The Kranz syndrome in the Gramineae as indicated by carbon isotopic ration. American Journal of Botany, 60: 505-513. DOI:10.1002/j.1537-2197.1973.tb05951.x |
Voznesenskaya E V, Franceschi V R, Kiirats O, et al. 2001. Kranz anatomy is not essential for terrestrial C4 plant photosynthesis. Nature, 414: 543-546. DOI:10.1038/35107073 |