 2007, Vol. 43
2007, Vol. 43文章信息
- 柴春山, 蔡国军, 孟少童, 陈炜青, 薛睿.
- Chai Chunshan, Cai Guojun, Meng Shaotong, Chen Weiqing, Xue Rui.
- 树苜蓿在甘肃的育苗试验
- Breeding Trial of Chamaecytisus palmensis in Gansu Province
- 林业科学, 2007, 43(12): 149-155.
- Scientia Silvae Sinicae, 2007, 43(12): 149-155.
-
文章历史
- 收稿日期:2007-02-05
-
作者相关文章


树苜蓿(Chamaecytisus palmensis)又称灌木苜蓿或苜蓿树(Tree Lucerne,Tagasaste),起源于非洲西北海岸大西洋的加拿利群岛(Canary Islands)的拉帕尔马岛(La Palma)上(Webb et al., 1985),是一种速生常绿豆科灌木(Getnet,1998),分布于拉帕尔马岛的亚热带、地中海式生物气候带(Javier et al., 2004),后从拉帕尔马岛传播到其他岛屿(Francisco-Ortega et al., 1993)。目前,智利、澳大利亚、新西兰、夏威夷、爪哇和东非等许多国家和地区有引种栽培(Aronson et al., 2002; Virtue et al., 2003),尤其是澳大利亚和新西兰现已广泛栽培且有百年历史(Snook,1996)。经世界各国对大量物种对比研究,认为树苜蓿在饲用、农林复合、防护、固土、降低水位、提高土壤肥力和促进退化土地恢复及园林绿化等方面具有很大的发展潜力(Getnet,1998; Aronson et al., 2002; Snook,1996; Lefroy et al., 1992; 2001)。甘肃于2002年5月首次将树苜蓿从澳大利亚新南威尔市州的Nerriga引进中国,并进行了种子催芽、育苗、造林、越冬等试验研究,部分内容作了初步报道(彭鸿嘉等,2003; 柴春山等,2004; 蔡国军等,2005)。现将2002—2005年的研究结果进行综合报道,以期为树苜蓿在中国的引种栽培提供科学依据。
1 植物学特征和生物学特性树苜蓿,属豆科(Leguminosae)蝶形花亚科(Papilionoideae),常绿灌木,喜光,速生,1年生苗高约1~3 m,成株后达4~7 m,第3年开花,树形高大疏散,寿命长20~40 a。枝条细长,被有一层银灰色短柔毛。三出复叶,叶背密被茸毛,叶面光滑,嫩叶呈椭圆形或倒披针形。冬、春两季开花,蝶形花冠,花白色,3~8朵为1束,单体雄蕊,自花受粉。荚果细长,具短茸毛。种子光滑呈黑色,卵圆形,具一突起的种翅。适于降水量多、无霜冻、排水好的亚热带和暖温带地区生长。能适应多种土壤类型和土壤肥力,最适pH值6.0~7.0的微酸性土壤,不适应积水土和碱性土。植株不耐荫,不耐霜冻,尤其是幼苗,这一敏感性能限制其成功地建植。固氮作用显著,能提高土壤肥力和促进退化土地的恢复与植物生长。茎、叶蛋白质含量高,嗜口性好,是一种灌木饲料。
2 试验地概况试验地1为定西市水土保持研究所苗圃(104°23′ E,35°21′ N),海拔1 900 m,属温带大陆性季风气候,年均气温6.3 ℃,极端高温34.4 ℃,极端低温-27.1 ℃,年均降水量425 mm,无霜期140 d,土壤为黄绵土,肥力中等,pH值8.0左右,有灌溉条件;试验地2为天水市中滩镇三阳苗圃(105°53′ E,34°33′ N),海拔1 084.2 m,属北暖温带半湿润气候,年均气温11.1 ℃,极端高温37.2 ℃,极端低温-17.6 ℃,年均降水量496. 5 mm,土壤为黄绵土,肥力良好,有灌溉条件;试验地3为陇南市武都区青家坝苗圃(104 ° 55′ E,33°24′ N),海拔1 081.7 m,属北亚热带半湿润季风气候,年均气温14. 9 ℃,极端高温39.0 ℃,极端低温-8.6 ℃,年均降水量474 mm,土壤为石灰性褐土,肥力较差,pH值8.0左右,有灌溉条件。
3 材料与方法 3.1 种子催芽、发芽试验及生活力测定催芽试验采用砂纸磨擦、浓硫酸浸种、碳酸钠浸种、洗衣粉浸种、沸水浸种和变温处理6种方法(彭鸿嘉等,2003),以清水浸泡为对照,以处理浸泡种子的吸胀率分析不同方法的催芽效果(彭鸿嘉等,2003);催芽种子生活力测定采用靛蓝染色法(孙时轩,1995);发芽试验采用实验室常规发芽试验,种子是经6种方法催芽处理的吸胀种子。
3.2 育苗种子处理育苗种子采用洗衣粉加热水处理(蔡国军等,2005)。由于树苜蓿种子存在硬实现象,洗衣粉浸泡处理无法得到一致的种子,所以经处理的种子分3类:一是吸胀并露白的种子;二是吸胀未露白的种子;三是未吸胀处于硬粒状态的种子。
3.3 容器育苗于2002—2005年在3个试验地进行。营养土用圃土、蛭石、河沙、肥料(农家肥和化肥)按5:3:2:1的比例配制(蔡国军等,2005),营养袋规格为6 cm×17 cm直筒无底塑料袋和8 cm×14 cm的营养钵。播种量每袋3~5粒,覆土厚度2~3 cm。播后进行洒水、除草管理。出苗后,在整个苗床随机选取3畦,每畦按前、中、后3个部位调查出苗率和成活率。
3.4 扦插育苗于2003年7月31日和9月9日在定西市水土保持研究所苗圃进行。第1次采用简易塑料大棚,苗床设在棚口,人工洒水;第2次采用普通日光温室,苗床设在靠中间部位,自动微喷管理。插穗采自定西当年4月所育容器苗主干的上、中、下3个部位(柴春山等,2004)。均用50 mg·kg-1的ABT生根粉浸泡处理,第1次处理2 h,第2次处理10 h,设清水浸泡为对照。苗床下层为圃土,上层为厚5 cm的细沙。扦插株行距5 cm×5 cm,深度5 cm。试验采用完全随机区组设计,每个处理3次重复。扦插1周后进行新叶、愈伤组织和生根观测,待苗生根成活后,进行愈伤率和生根率调查。
3.5 直播育苗于2002—2005年在3个试验地进行。用硫酸亚铁进行土壤消毒和调节pH值。采用条播,条间距30 cm,覆土厚度3 cm,播后进行洒水、除草及松土管护。2002年6月在定西进行不同覆土厚度(1~2 cm、2~3 cm和4~5 cm)和不同覆盖(覆膜、覆草和无覆盖)播种试验,种子为吸胀未露白种子。覆土试验在定西水保所苗圃进行,条播,条间距30 cm,株间距约2 cm,每个处理3次重复,播后进行洒水、除草及松土管护;覆盖试验在定西安家沟流域阴坡梯田地进行,穴播,穴间距1 m×1 m,覆土厚2 cm,每穴6粒,每个处理4次重复,播后不进行任何管护,覆膜分塑料膜有孔和无孔两种。覆土试验每个重复随机选取1畦,每畦按前、中、后3个部位设1 m2样方,调查出苗率;覆盖试验每种处理随机选取24穴调查出苗率。
3.6 生长量测定定西2003年所育容器苗调查采用定时定株测定方法,在整个苗床随机抽取70株苗为固定样本进行编号供测定,测定季节为5—11月,每10 d测定1次;其他容器苗、直播苗和扦插苗在10月底调查,调查样本为55株。
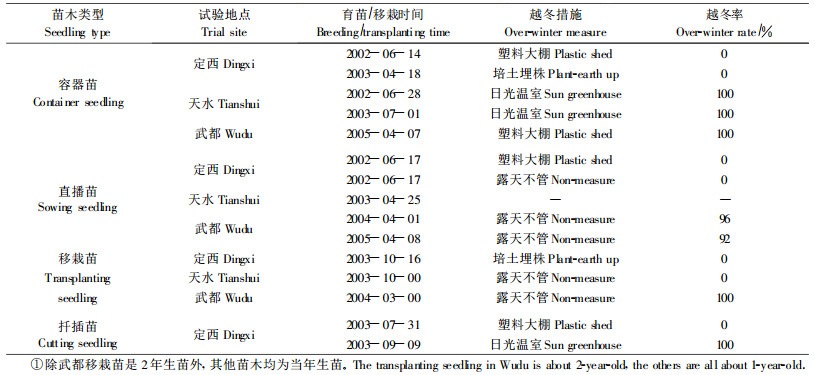
3.7 越冬试验越冬试验采取搭盖塑料大棚、日光温室、培土埋株和露天等4种方式,在3个试验地进行。4种越冬方式都是先灌水,后搭棚或培土,依各越冬地气候状况在每年10月下旬至11月上旬进行,第2年春季苗木返青后进行越冬率调查。
4 结果与分析 4.1 种子测定和催芽试验物理指标测定表明,树苜蓿成熟种子光滑呈黑色,未成熟或尚未成熟种子呈淡绿色或红褐色,长约4 mm,手感滑腻,千粒质量23.92 g,净度93.17%,硬实率63.0%。催芽试验结果表明,6种催芽方法存在明显差异。其中,砂纸磨擦效果最好,其次是浓硫酸和变温催芽,碳酸钠和洗衣粉效果居中,沸水浸种效果相对较差(彭鸿嘉等,2003)。6种催芽方法种子平均吸胀率为31.9%,明显高于对照(17%),说明催芽处理对去除种子表面油脂和软化种皮是有效的(彭鸿嘉等,2003)。在大规模育苗时,考虑到催芽方法的安全、简便和有效性,采用沸水或热水加洗衣粉的催芽方法。
4.2 催芽种子生活力测定结果表明,6种方法催芽后的种子生活力较高,平均达82%,6种催芽方法没有明显影响种子的活力,但随着浓硫酸和洗衣粉处理时间的延长,种子活力开始下降,由原来的90%分别降为85%和75%(彭鸿嘉等,2003),说明用以上6种方法进行催芽处理时要掌握适宜的处理浓度和处理时间。
4.3 催芽种子发芽试验经6种方法催芽吸胀的种子室内发芽率高达94.5%,发芽速度快,第2天就开始发芽,第3天达到高峰期,6天内79%的种子发芽,10天基本完成出苗。说明树苜蓿种子虽发芽困难和缓慢,但只要进行适当的催芽处理并选用吸胀的种子育苗,种子不但发芽率高而且发芽速度快。
4.4 容器和直播育苗出苗观测从表 1可知,树苜蓿种子出苗期约2~16 d,齐苗期4~13 d,3类种子出苗期和齐苗期具有明显差别,泡胀露白种子播后需2~3 d出苗,4 d出齐,出苗最早、齐苗最快;泡胀未露白种子需4~9 d出苗,9~10 d出齐,出苗和齐苗时间和速度居中;硬粒种子需13~16 d出苗,9~13 d出齐,出苗期最长,需在土壤中吸足水分才能萌发。试验中直播苗比容器苗出苗和齐苗略晚几天,出苗率差别不明显,但成活率因土壤板结和病菌感染比容器苗低。3类种子平均出苗率和成活率分别是91.1%和82. 1%,未泡胀硬粒种子出苗率最好,平均达95.4%,但成活率最差,平均为76.6%;其次是泡胀露白种子,平均出苗率和成活率为94.5%和88.0%;泡胀未露白种子出苗率和成活率较差,平均为85.0%和84.7%。
|
|
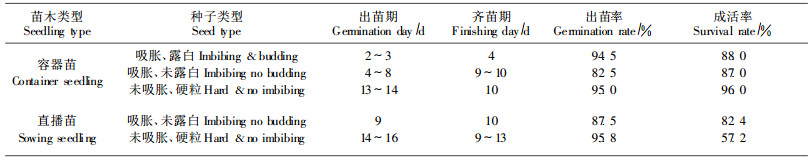
1) 覆土厚度育苗试验3种不同覆土厚度的出苗期和齐苗期相差不大,1~2 cm和2~3 cm的出苗期和齐苗期相同,均为9 d和10 d,4~5 cm的出苗期略晚2 d(表 2)。但3种不同覆土厚度的出苗率差别非常明显,2~3 cm出苗率最好(9 2%),其次是1~2 cm,4~5 cm出苗率极差,说明树苜蓿最适宜的覆土厚度为2~3 cm。
|
|

2) 覆盖播种育苗试验由表 3可见,覆盖和对照齐苗期相差不大,为9~12 d;出苗期有差异,覆盖比对照早3 d;出苗率差异较明显,覆盖平均出苗率为82%,明显高于对照(51%)。说明覆盖处理能提高树苜蓿大田直播育苗的出苗率和出苗速度,尤其在春季采用塑料薄膜覆盖既能提高地温,又能保持湿度,效果好,这一点在武都直播育苗中得到了证实。
|
|
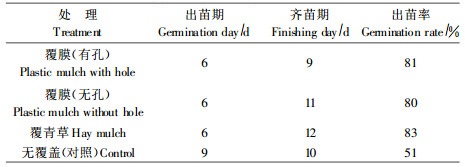
1) 扦插苗新叶、愈伤组织和生根观测从2次扦插结果看(表 4),树苜蓿扦插可以产生愈伤组织并能生根。扦插后约7~9 d长出新叶,13~15 d产生愈伤组织,20~26 d生根,平均愈伤率为54.2%~65.7%,平均生根率为11.3%~29.7%,发芽、生根快,愈伤率高,但实际生根率低,2次扦插平均生根率只有20.5%。9月扦插苗新叶萌发、愈伤组织产生和生根均比7月早,愈伤率和生根率高,其原因是9月比7月插条木质化程度高、ABT处理时间长和管理条件好。
|
|

2) 环境和管理条件对插穗愈伤组织和生根影响从表 4可知,2次扦插苗愈伤率较高,分别达54.2%(第1次)和65.7%(第2次),表明树苜蓿生根潜力较大。但2次实际生根率只有11.3%和29.7%,分别约有43%和36%的愈伤苗未生根,且第1次愈伤苗未生根率比第2次高,其原因与2次扦插的环境和管理条件有关(柴春山等,2004)。
3) 不同部位插穗对生根影响从插穗部位来看,插条上部和中部比下部生根率高(表 5)。7月份大棚扦插苗上、中、下的平均生根率分别是13.6%、11.2%和9.0%;9月份温室扦插苗上、中、下生根率分别是33.5%、31.2%和24.3%。可见,上部插穗生根率最高,中部次之,下部最低,说明越靠枝条的上部越易生根。
|
|
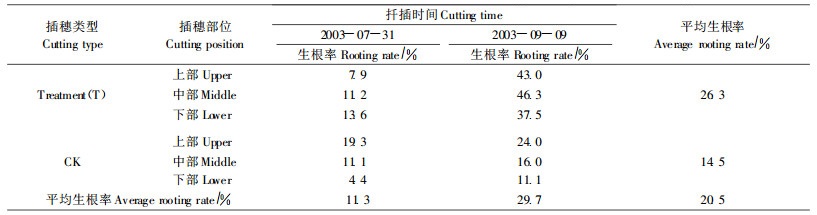
4) ABT处理对插穗生根影响ABT处理苗(T)生根率比对照(CK)高,平均生根率分别为26.3%和14.5%(表 5),ABT处理效果较明显。第2次T平均生根率(41.7%)明显高于CK (17.3%),第1次T(10.9%)和CK(11.6%)差异不大,两次产生差异的原因有可能是AB T处理时间不同所致。
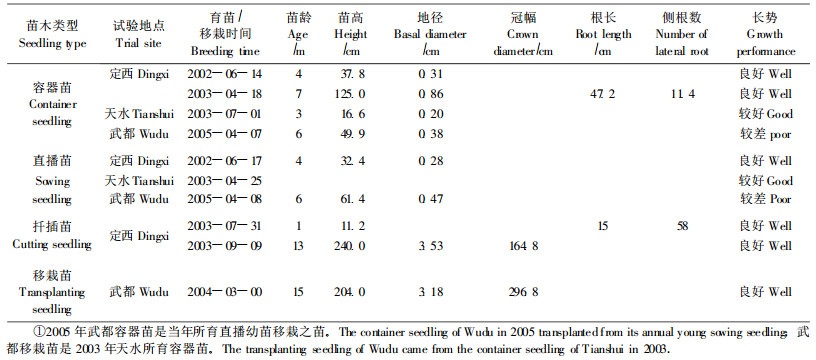
4.7 生长量测定1) 容器苗生长量容器苗3—7月龄平均高16.6~125.0 cm,地径0.20~0.86 cm(表 6)。其中2003年定西7月龄苗平均高>1 m,天水3月龄平均高达16.6 cm,说明树苜蓿容器苗高生长迅速,但平均地径不到1 cm(因容器苗未移植)。播种月份相同、地点不同、生长期接近的容器苗相比,定西生长优于武都(表 6)。但容器苗根系(5月龄)因未移植而生长较差,平均根长只有47.2 cm,远小于苗高生长。从长势看,1年生容器苗定西长势好,天水次之,武都较差。定西容器苗在1个年生长周期内苗高和地径生长曲线都呈“S"形(图 1),即具有1次迅速生长期和2次缓慢生长期,速生期主要在7—9月;地径生长曲线比苗高平缓,地径速生期开始时间比苗高滞后,说明地径比苗高增长慢。
|
|

|
图 1 树苜蓿苗高和地径生长曲线 Fig. 1 The growth curve of tree height and basal diameter of C. palmensis |
2) 直播苗生长量直播苗4—6月龄平均高32.4~61.4 cm,平均地径0.28~0.47 cm(表 6)。直播苗在3个育苗地当年生平均高均不到1 m,地径不到0.5 cm,说明树苜蓿直播苗比容器苗生长慢,这主要与两者的土壤养分和土壤疏松状况不同有关。就同一地点、同一时间播种的容器苗和直播苗相比,定西容器苗比直播苗生长快(2002年6月),而武都则相反(2005年4月),这与武都容器苗是当年所育直播苗移栽而来从而延缓生长期有关。
3) 移栽苗和扦插苗生长量不足2年生移栽苗和扦插苗平均高>2 m,平均地径>3 cm,平均冠幅将近3 m(表 6),表现出明显的速生特性。尤其是扦插苗,与容器苗相比,由于根系生长没有容器袋等限制,加之苗床疏松的良好生长环境,生长非常快,扦插生根后1个月新梢平均长11.2 cm,平均根长15 cm,平均侧根数58个,比定西5月龄容器苗侧根数多而明显;与移栽苗相比,由于没有缓苗期,苗高和地径增长比月龄大的武都移栽苗还快。
4.8 越冬试验试验结果表明(表 7),在定西和天水,凡是日光温室越冬当年生容器苗和扦插苗,即便育苗时间比其他越冬苗晚1—3个月,但越冬率为100%;采取培土埋株和搭塑料大棚保护措施的苗木,尽管育苗时间早,但越冬率为0。在武都,气温低于- 5.0 ℃时,露天越冬当年生直播苗地上部分和2年生移栽苗新梢部分发生冻害,但第2年春均能恢复,苗木能安全越冬,平均越冬率达94%;采取只在夜间搭盖塑料棚而白天敞露的当年生容器苗(2005年),没有发生冻害现象,越冬率为100%。由此看来,树苜蓿当年生苗在武都基本能自然越冬,在定西和天水不能。同时,试验还发现,1年生以上苗木在武都没有任何越冬障碍,说明随苗龄的增加,树苜蓿抗寒性增强。Getnet(1998)也指出树苜蓿当年生幼株冬季发生冻害但第2年植株包括幼嫩再生枝条变得抗寒、抗旱且能忍耐-7.7 ℃的低温。
|
|
树苜蓿种子具有油脂和硬实现象,种子发芽缓慢,播前必需进行催芽处理。本研究6种催芽方法中,碳酸钠和洗衣粉催芽效果虽然居中(种子吸胀率27.9%),但此方法比较简单、安全和实用,在大规模育苗中比较适宜采用。
树苜蓿经催芽的3类种子中,泡胀露白种子出苗最早且齐苗最快;泡胀未露白种子出苗速度居中,但出苗率最差;硬粒种子出苗期最长,需半个月,但出苗率最好。泡胀未露白种子出苗率最差是由于其含水量高,播后不能象吸胀露白种子那样快速出苗,加之育苗时高温、高湿致使种子大量霉烂,从而造成出苗率较低。相比之下,处于硬粒状态的种子出苗率和成活率较好,这可能是其由于没露芽和未吸胀水而未受到损伤或失水、腐烂的缘故,此外也是硬实种子活力高于非硬实种子的缘故(徐本美等,2006),说明树苜蓿大规模育苗种子只需除去表面油脂而不必待到种子吸胀水时播种更好。
树苜蓿扦插育苗可以产生愈伤组织并能生根,平均愈伤率和生根率为60.0%和20.5%。由于条件限制,本研究只做了低浓度ABT处理的小规模扦插试验,今后有必要进行ABT高浓度及多种激素处理的大规模扦插试验,以期提高生根率。
树苜蓿是阳性速生树种,当年生苗平均高>1.0 m,地径接近1.0 cm;不足2年生苗平均高>2 m,地径>3 cm,冠幅接近3 m。扦插苗生长最快,长势好;容器苗居中;直播苗较差。但由于所育3类苗木当年都未经移栽,密度大,严重限制了其地径、根系和冠幅的生长发育,致使植株生长细弱,在一定程度上影响了当年生苗木的生长量。但经对比研究,树苜蓿不论是容器苗还是扦插苗,与定西同期所育沙棘(Hippophae rhamnoides)、柠条(Caragana microphylla)和甘檬柽柳(Tamarix austrmongolica)的同类苗木在苗高、地径、侧枝以及冠幅方面相比,生长量依然远大于后者,具有很好的速生特性和较大的生物量,这与国外报道结论相一致(Aronson et al., 2002)。
树苜蓿当年生苗在武都尚能自然越冬但生长较差,在定西和天水生长良好但不能自然越冬,这主要与3个试验地气候尤其是气温有着直接关系。受自然选择的影响,各树种与其自然分布区生态、环境有着天然的适应,与原产地相似的环境条件下最有可能发挥其生物学特性(Wright,1978)。树苜蓿原产地加拿利群岛属于亚热带地中海式气候,常年气温保持在20~ 25 ℃左右。甘肃引种树苜蓿种源地Nerriga介于亚热带和暖温带的冬暖性气候带,极端低温只有-9.0 ℃,极端高温虽达39.7 ℃,但1年中≥35 ℃的平均天数不超过5 d(1898—20 04年),气候与原产地相差不大。相比之下,甘肃引种试验地气候与原产地和种源地相差悬殊,虽然武都极端最低气温只有-8.6 ℃,接近于种源地,但≥35 ℃高温持续时间长,不利苗木生长,而定西和天水夏季温度相对武都较低且持续时间短,有利苗木生长,但冬季气温过低,最低气温均在-10.0 ℃以下,定西甚至达-27.0 ℃,不利苗木越冬。中国和澳大利亚两国气候分类系统不兼容,在澳大利亚被认为是温带的大部分地区相当于中国的亚热带地区,造成气候匹配的失准,进而导致引种决策的失误和引种时冻害的发生(阎洪,2006)。在影响外来树种引种成败的环境诸因子中,气候因素具有决定性的作用(Woodward,1987)。除气候因素外,育苗时间晚,苗木未移栽或移栽迟,造成各类苗木根系不发达和木质化程度低,也在一定程度上影响了树苜蓿苗木越冬。
考虑到树苜蓿是一种饲用价值较高的灌木树种,具有适宜山区栽植和抗旱的特性,并在世界上许多引种国家作为饲用、薪柴和农林复合树种在丘陵地区普遍栽培(Lefroy et al., 1992;Niang et al., 1996;Getnet,1998;Aronson et al., 2002),甘肃省林业科学研究院将此树种作为饲用和荒山造林树种引进到甘肃中部的干旱地区定西,但经过1年的试验研究发现,即便采取越冬保护,苗木也无法安全越冬,当年所育几千株苗木全部冻死,致使试验地转移到天水,同时保留定西试验点,但第2年2个试验点的苗木除日光温室外,近万株其他方式越冬的各类苗木又全军覆没,又一次将试验点转移到陇南的武都。尽管当年生苗木在武都能安全越冬,但也存在需夜间保护的问题,大面积栽培是不可行的,加之夏季气温过高,生长状况远不如定西和天水。所以,无论从树苜蓿最初的引种意愿还是实际的引种结果来看,它在甘肃的引种是失败的,教训也是深刻的。树种在跨国间的引种除了考虑宏观的气候因素外,还要考虑微观的气候因子、多年的气象资料和不同国家的气候分类系统,如从澳大利亚引种树苜蓿不能单从气候类型考虑,因为澳大利亚仅温带气候类型就包括3种雨型,树苜蓿主要分布在冬雨型,而甘肃几乎是夏雨型。即便气候类型相同,也因分类系统的不同而大不相同,所以引种发生冻害甚至失败是不可避免的。
通过对树苜蓿在定西、天水和武都3个不同纬度区的生长表现和越冬状况试验研究并综合分析甘肃、中国和树苜蓿原产地、种源地及世界上其他引种栽培区的气候特征,并根据气候相似原理,从越冬的气候角度考虑,初步认为树苜蓿在甘肃陇南纬度33°24′及以南地区可以试种;在中国,中亚热带、南亚热带和热带地区气候比较接近澳大利亚树苜蓿分布区,可能适宜其引种栽培,但有待试验研究。
蔡国军, 柴春山, 孟少童, 等. 2005. 引进优良饲料灌木树苜蓿容器育苗试验初报. 草业科学, 22(10): 55-58. DOI:10.3969/j.issn.1001-0629.2005.10.012 |
柴春山, 蔡国军. 2004. 引进优良饲料灌木树苜蓿嫩枝扦插育苗试验初报. 甘肃林业科技, 29(4): 5-8. DOI:10.3969/j.issn.1006-0960.2004.04.002 |
彭鸿嘉, 蔡国军, 柴春山, 等. 2003. 优良引进饲草灌木树苜蓿种子的6种催芽试验. 甘肃林业科技, 28(3): 5-8. |
孙时轩. 1995. 造林学. 2版. 北京: 中国林业出版社.
|
徐本美, 孙运涛, 李锐丽, 等. 2006. 舞草硬实种子的高活力性状研究. 林业科学, 42(10): 54-58. |
阎洪. 2006. 中国和澳大利亚的气候比较研究. 林业科学, 42(8): 30-36. |
Aronson J, Ovalle C, Avendaño J, et al. 2002. Agroforestry tree selection in central Chile: biological nitrogen fixation and early plant growth in six dryland species. Agroforestry Systems, 56(2): 155-166. DOI:10.1023/A:1021345318008 |
Francisco-Ortega J, Newbury H J, Ford-Lloyd B V. 1993. Numerical analyses of RAPD data highlight the origin of cultivated tagasaste (Chamaecytisus proliferus ssp.palmensis) in the Canary Islands. Theor Appl Genet, 87: 264-270. DOI:10.1007/BF00223775 |
Getnet A. 1998. Biomass yield, botanical fractions and quality of tagasaste(Chamaecytisus palmensis) as affected by harvesting interval in the highlands of Ethiopia. Agroforestry Systems, 42(1): 13-23. DOI:10.1023/A:1006099521986 |
Javier D C, Milagros L B, Ricardo P G. 2004. Screening for plant growth-promoting rhizobacteria in Chamaecytisus proliferus (tagasaste), a forage tree-shrub legume endemic to the Canary Islands. Plant and Soil, 266: 261-272. |
Lefroy E C, Dann P R, Wildin J H, et al. 1992. Trees and shrubs as sources of fodder in Australia. Agroforestry Systems, 20(2): 117-139. |
Lefroy E C, Stirzaker R J, Pate J S. 2001. The influence of tagasaste (Chamaecytisus proliferus Link.) trees on the water balance of an alley cropping system on deep sand in south-western Australia. Aust J Agric Res, 52: 235-246. DOI:10.1071/AR00035 |
Niang A, Ugiziwe J, Styger E, et al. 1996. Forage potential of eight woody species: intake and growth rate of local young goats in the highland region of Rwanda. Agroforestry Systems, 34(2): 171-178. DOI:10.1007/BF00148160 |
Snook L C. 1996. Tagasaste: A productive browse shrub for sustainable agriculture. Washingtone DC:: Natiunal Academy Press.
|
Virtue J G, Melland R L. 2003. The Environmental weed risk of revegetation and forestry plants. Report DWLBC: 93-98. |
Webb C J, Shand E. 1985. Reproductive biology of tree Lucerne (Chamaecytisus palmensis, Leguminosae). New Zealand Journal of Botany, 23: 597-606. DOI:10.1080/0028825X.1985.10434230 |
Woodward F I. 1987. Climate and plant distribution 1: Vegetation and climate. Cambridge University Press.
|
Wright J W. 1978. Introduction of forest genetics. MA, USA: Academic Press.
|