 2007, Vol. 43
2007, Vol. 43文章信息
- 冯立国, 邵大伟, 生利霞, 赵兰勇, 何小弟.
- Feng Liguo, Shao Dawei, Sheng Lixia, Zhao Lanyong, He Xiaodi.
- 野生玫瑰的花粉形态及其起源与演化的探讨
- Pollen Morphology of Wild Rosa rugosa and Discussion on Its Origin and Evolution
- 林业科学, 2007, 43(12): 76-80.
- Scientia Silvae Sinicae, 2007, 43(12): 76-80.
-
文章历史
- 收稿日期:2007-04-02
-
作者相关文章



2. 山东农业大学林学院 泰安 271018
2. College of Forestry, Shandong Agricultural University Tai'an 271018
花粉形态特征是植物在长期的进化过程中不断演化和发展形成的, 其中带有大量有关演化的信息, 它是由基因控制的, 受外界环境条件的影响很小, 具有很强的遗传保守性和稳定性, 是探讨植物起源、演化及亲缘关系的重要依据之一(王开发等, 1983; 汪祖华等, 1990; Erdtman, 1978)。野生玫瑰(Rosa rugosa)是国家二级濒危植物, 在我国天然分布于吉林图们江河口、辽宁南部海岸以及山东东部沿海海岸(傅立国等, 1992), 其基因组中包含芳香、抗寒、抗黑斑病等优良性状基因, 是栽培玫瑰以及蔷薇属花卉育种的重要种质资源。长期以来, 国内对野生玫瑰的研究主要侧重于生态学调查、繁殖栽培技术以及保护策略等方面(陈建军等, 1997; 张淑萍等, 2005; 刘海峰等, 2006), 有关中国野生玫瑰花粉形态的研究国内外尚未见报道。本文在对国内野生玫瑰种质资源进行全面调查的基础上, 以原产地和形态特征均具代表性的6个野生玫瑰类型为研究对象, 在扫描电镜下系统地观察、比较了其花粉形态, 并进行了聚类分析, 在此基础上分析了其亲缘关系并对中国野生玫瑰的起源与演化进行了探讨, 旨在为野生玫瑰系统学及保护生物学研究提供孢粉学资料。
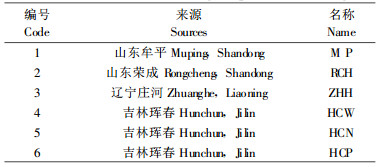
1 材料与方法 1.1 试验材料试验于2006年5—8月进行, 6份花粉材料分别采自山东、辽宁和吉林省的野生玫瑰居群(表 1)。选择形态特征具有代表性的居群, 从饱满、即将开放的花蕾中直接取花药, 置于硫酸纸袋中, 在室内自然干燥。
|
|
将经自然干燥后的供试花粉均匀涂布于粘有双面胶带的样品台上, 于IB-5型粒子溅射仪中喷金, 然后置于日立S-570扫描电镜下观察记录, 并选取有代表性的视野分500×(群体)、2 000×(个体, 赤道面)、2 500×(个体, 极面)和12 000×(局部, 赤道面中央区)进行拍摄(见图版), 在数码照片上利用CAD软件测量正常花粉粒的极轴长P、赤道轴长E、孔穴直径D、条嵴宽W、条嵴间距、条嵴密度以及孔穴密度, 观察记录各花粉的赤道面观、极面观以及两极的形状, 萌发器官及外壁纹饰特点, 每种花粉测量20粒, 求平均值, 并计算标准差。以(P×E)表示花粉粒大小, P/E值表示花粉粒的形状(P/E>2表示花粉粒为超长球形, P/E为1.14 ~ 2表示花粉粒为长球形)(王开发等, 1983)。
1.3 统计分析以供试的6份花粉作为分类运算单位, 根据上述观测结果, 按照数量分类学性状选择的原则, 剔除不变性状和逻辑相关性状, 共选取9个性状, 其中6个为数值性状, 即P、P/E、D、D/W、嵴间距及孔穴密度, 3个为无序多态性状, 即赤道面观、萌发器官及外壁纹饰特点。性状量化方法为数值性状直接取其数值, 无序多态性状则采用分解法进行编码(徐克学, 1994)。将数据输入计算机后标准化, 采用SAS软件进行聚类分析, 聚类单元之间的遗传距离系数采用马氏距离系数, 选用聚类效果较好的UPGMA法。
2 结果与分析 2.1 花粉形态野生玫瑰的花粉粒为长球形或超长球形, MP的花粉粒最大, ZHH、RCH次之, 来自吉林珲春的HCW、HCN和HCP的花粉粒相对较小。MP花粉粒表面的条嵴宽而稀疏, 嵴间距最大, 嵴密度最小, 而HCP花粉粒表面的条嵴窄而密, 嵴间距最小, 嵴密度最大, 二者相差几乎1倍(表 2)。
|
|
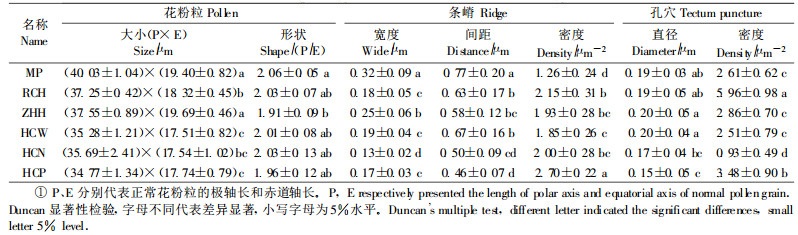
野生玫瑰花粉粒表面均分布有大小、疏密不等的孔穴, 部分较大的形成穿孔, 未形成穿孔的则呈小穴状(图 1)。HCN和HCP花粉粒表面的孔穴直径较小, 其余野生玫瑰之间差异不大; 孔穴密度以RCH花粉为最大, 达到了5.96个·μm-2, 而HCN花粉表面仅有0.93个·μm-2, 相差近7倍(表 2)。

|
图 1 野生玫瑰的花粉形态及其起源与演化的探讨 Fig. 1 Pollen morphology of wild Rosa rugosa and discussion on its origin and evolution |
野生玫瑰花粉极面观均呈三裂圆形, 赤道面观MP、RCH和ZHH呈长椭圆形偏矩形, HCW和HCN呈长椭圆形, 而HCP呈椭圆形偏纺锤形; MP、RCH和ZHH花粉的两极形状均为平截形, 而来自吉林珲春的HCW、HCN和HCP花粉的两极均呈圆弧形(图 1)。
2.2 花粉外壁纹饰6个野生玫瑰居群花粉的外壁均具条嵴, 嵴洼处散生有覆盖层穿孔, 属于条纹-穴状纹饰, 但条嵴及穿孔的形态和分布差异明显(图 1)。
MP :花粉外壁分布有深浅不一、形状各异的嵴洼, 嵴洼内散生有大小不一、形状不规则的孔穴, 有近一半形成穿孔。条嵴宽而疏, 较平坦, 有分叉, 走向不规则。
RCH :花粉表面密布小孔穴, 圆形或近圆形, 基本都形成穿孔, 位于高高隆起的网嵴之间, 两极端无孔穴。条嵴窄而密, 分叉较多, 由赤道面一端纵向平行渐转为斜向平行进而呈弧状平行, 至另一端转为斜向平行。
ZHH :花粉粒表面密布小孔穴, 大小不一, 形状不规则, 在中央区分布的孔穴较稀疏, 浅而小, 大部分未形成穿孔, 呈浅小穴状, 而近两极端的孔穴较大, 基本都形成穿孔。条嵴较宽, 较平坦, 由赤道面一端斜向平行, 至中部转为纵向平行, 至另一端又转为斜向平行。
HCW:花粉粒表面密布小孔穴, 大多数都形成穿孔, 穿孔形状不规则, 未形成穿孔的小穴形状较规则, 圆形或近圆形, 两极端孔穴分布较稀疏, 部分区域甚至无孔穴。条嵴平坦, 隆起不明显, 走向不规则。
HCN :花粉粒表面稀疏的分布有小孔穴, 孔穴近圆形, 小而浅, 基本都未形成穿孔, 两极端几乎无孔穴。条嵴隆起不明显, 窄, 较密, 有分叉, 斜向平行。
HCP :花粉粒表面密布圆形或近圆形的小孔穴, 约2/3形成穿孔, 位于高高隆起的网嵴之间。条嵴窄而密, 间距均匀, 有分叉, 在赤道面中部纵向平行, 进而转为斜向平行, 近极端呈弯曲弧状平行。
2.3 花粉的萌发器官6个野生玫瑰居群的花粉均具3条孔沟, 沿极轴方向以等间距环状分布, 按照Erdtman(1978)的NPC分类系统, 属于N3P4C5型花粉。从花粉赤道面可见1 ~ 2条萌发沟, 极面可观察到3条, 沟长几达两极。HCN花粉的萌发沟不闭合, 在沟端成约30 °夹角或有所扩展, 其余野生玫瑰花粉的萌发沟均呈闭合状, 不开裂, 未在沟端形成夹角(图 1)。
2.4 聚类分析如图 2所示, HCW和HCN之间的遗传距离系数最小, 因而最早聚在一起; MP首先与ZHH聚在一起, 然后二者又与RCH聚在一起; 而HCP与另外5个野生玫瑰的遗传距离系数最大, 因而聚合最晚。

|
图 2 野生玫瑰花粉性状的聚类分析 Fig. 2 Dendrogram of the wild Rosa rugosa by clustering analysis according to the pollen characteristics |
由等级结合线Lq1=15.49可将6个野生居群分为2组, 第1组包括MP、ZHH、RCH、HCW和HCN, 而HCP单独划为1类; 进一步做等级结合线Lq2=11.25可将6个野生居群分为3组, 第1组包括MP、ZHH和RCH, 第2组包括HCW和HCN, 而HCP仍然单独划为1组。
3 讨论 3.1 野生玫瑰花粉形态的多样性及分类意义扫描电镜下, 6个野生玫瑰居群的花粉性状表现出丰富的多样性, 尤其是在外壁纹饰上差异明显, 突出表现在条嵴走向、深浅、宽度、密度以及孔穴密度等方面。花粉粒大小、赤道面观以及两极形状也存在较大的差异。花粉外壁纹饰是进行植物分类的重要的、最有价值的依据之一(朱岭仁等, 1995)。野生玫瑰花粉的外壁纹饰形态各异, 6个野生居群完全不同, 表明这些纹饰特征具有重要的分类价值, 可以作为野生玫瑰分类、鉴定的重要依据。
3.2 野生玫瑰的亲缘关系野外调查发现, MP、ZHH、RCH和HCW的形态特征较为接近, 只是在小叶形状、花色上有区别, 但差别较小, 部分性状甚至在不同的居群中都出现, 单通过植株外部形态特征很难确定它们之间的亲缘关系。花粉性状的聚类分析结果表明, MP与ZHH的遗传距离较近, 聚合较早, 然后二者又与RCH聚在一起(图 2), 说明三者具有一定的亲缘关系, MP与ZHH的亲缘关系较近。
吉林珲春地区野生玫瑰的形态变异甚为丰富。HCW的小叶呈宽椭圆形, 先端急尖, HCN的小叶形态与HCW不同, 其小叶狭长, 先端尖, 基部宽楔形, 但二者除了叶部特征存在差异外, 其他形态特征非常相似, 如花均为紫红色, 单瓣, 枝条灰白色, 皮刺黄色, 多而密, 果实扁球形或近球形, 红色。花粉性状的聚类分析结果也表明二者的遗传距离最小, 亲缘关系密切。HCW和HCN随后在Lq=13.44处与MP、ZHH及RCH聚到一起(图 2), 说明它们之间也存在一定的亲缘关系。
HCP是我们在野外调查过程中发现的一个形态特殊的居群, 它与HCW的区别在于其花萼、叶轴及叶柄均呈红色, 老枝灰褐色, 皮刺多而密, 当年刚萌生新枝的枝刺红色, 多而密, 秋季落叶后枝干呈紫红色, 皮刺稀疏。植株外部形态特征表明HCP与其他野生玫瑰差别较大。本研究花粉性状的聚类分析结果表明, HCP与另外5个野生居群的遗传距离系数最大, 2次作等级结合线都单独划为1组(图 2), 说明HCP与其他5个野生居群的亲缘关系确实较远。
3.3 野生玫瑰花粉形态的演化及野生玫瑰起源的探讨Erdtman(1962)认为, “被子植物花粉外壁纹饰的演化是由无结构层(光滑)向穿孔(穴状)发展, 再由穿孔继续演化成条纹状类型”, 从他的论述中可以看出, 条嵴的有无、宽窄以及深浅等不仅反映花粉外壁纹饰的差异, 而且还可以作为衡量被子植物进化程度的参考之一。张秀英等(1997)对同为蔷薇科的桃(Prunus persica)的花粉进行扫描电镜观察后, 结合Erdtman的观点分析认为, 花粉外壁的条纹从分枝短且粗、排列不整齐向分枝长、排列整齐发展, 并且条嵴从宽、平向窄、嵴间沟深进化。本研究观察到6份野生玫瑰花粉的条嵴形态各异, MP花粉的条嵴最宽, 较平坦, 分叉短而粗, 排列不整齐; ZHH花粉的条嵴宽度仅次于MP, 较平坦, 条嵴及分叉排列整齐; HCW的条嵴明显窄、平坦且排列不整齐, 而RCH、HCN和HCP的条嵴更窄, 排列整齐, 且RCH和HCP的条嵴明显高高隆起, 嵴间沟深。这说明MP的花粉可能在进化上最原始, ZHH次之, HCW和HCN再次之, 而RCH和HCP则可能是最进化的类型。
关于花粉大小的演化顺序, 目前公认比较原始的被子植物花粉体积都是比较大的, 由大到小进化(周守标等, 2005)。本研究观察到MP的花粉粒最大, ZHH, RCH次之, 而HCN、HCW和HCP的花粉粒相对较小(表 2), 这说明吉林地区的野生玫瑰可能是较进化的居群, 山东和辽宁地区的野生玫瑰可能是较原始的居群, 而山东牟平的野生玫瑰则可能是最原始的居群。
综上所述, 中国野生玫瑰可能最初起源于山东东部沿海, 后经某种途径传播到辽宁南部沿海, 而后继续向北发展到吉林珲春地区。山东东部沿海的野生玫瑰是最原始的居群, 而荣成野生玫瑰可能是山东东部沿海居群中较进化的居群, 牟平野生玫瑰则是较原始的居群。在吉林珲春地区, HCW可能是相对较原始的居群, HCN由其演化而来, 而HCP则可能是最进化的居群。由于长期的地理隔离及居群隔离, 加之受当地独特的地质结构和气候条件的影响, 各地野生玫瑰在长期的进化过程中均发生了不同程度的变异, 遗传差异逐渐加大。
陈建军, 王景辉, 王东升, 等. 1997. 吉林珲春地区野生玫瑰的生态调查及栽培技术. 中国林副特产, (2): 52-53. |
傅立国, 金鉴明. 1992. 中国植物红皮书-稀有濒危植物. 第1册. 北京: 科学出版社.
|
刘海峰, 全炳武, 郭孟瑶, 等. 2006. 野生玫瑰扦插繁殖技术的研究. 延边大学农学学报, 28(2): 83-87. DOI:10.3969/j.issn.1004-7999.2006.02.002 |
汪祖华, 周建涛. 1990. 桃种质的亲缘演化关系研究:花粉形态分析. 园艺学报, 17(8): 161-168. |
王开发, 王宪曾. 1983. 孢粉学概论. 北京: 北京大学出版社.
|
徐克学. 1994. 数量分类学. 北京: 科学出版社.
|
张淑萍, 王仁卿, 杨继红, 等. 2005. 胶东海岸野生玫瑰(Rosa rugosa)的濒危现状与保护策略. 山东大学学报:理学版, 40(1): 112-118. |
张秀英, 王雁, 王桂萍. 1997. 桃花种质资源花粉形态的观察与比较. 北京林业大学学报, 19(2): 57-62. DOI:10.3321/j.issn:1000-1522.1997.02.009 |
周守标, 余本祺, 罗琦, 等. 2005. 石蒜属植物花粉形态及分类研究. 园艺学报, 32(5): 914-917. DOI:10.3321/j.issn:0513-353X.2005.05.035 |
朱岭仁, 孙京田. 1995. 山东蔷薇科植物花粉亚显微形态研究. 山东师范大学学报:自然科学版, 10(2): 192-196. |
Erdtman G, 1962.花粉形态与植物分类.王伏雄, 译.北京: 科学出版社
|
Erdtman G.1978.孢粉学手册.中科院植物研究所古植物研究室孢粉组, 译.北京: 科学出版社
|