 2007, Vol. 43
2007, Vol. 43文章信息
- 谢寅峰, 黄晗, 汤玉香, 周坚.
- Xie Yinfeng, Huang Han, Tang Yuxiang, Zhou Jian.
- 镧对汞胁迫下矢竹叶片生理反应的调节
- Regulating Effects of Lanthanum on Physiological Response of Pseudosasa japonica Leaf to Mercury Stress
- 林业科学, 2007, 43(12): 39-44.
- Scientia Silvae Sinicae, 2007, 43(12): 39-44.
-
文章历史
- 收稿日期:2007-05-08
-
作者相关文章


汞是毒性很强的重金属元素, 是重金属污染的重要来源。当植物体中汞的积累浓度达到一定范围后, 它就会破坏细胞的结构, 轻则使植物体内代谢过程发生紊乱, 生长发育受阻, 重则可造成植物枯萎, 甚至衰老死亡(母波等, 2007)。汞可直接与蛋白质(包括酶)、核酸等生物分子中的活性基团如巯基、磷酸基等结合, 或置换其中的阳离子使其结构与功能遭到破坏, 高浓度的汞也导致蛋白质的沉淀变性。植物叶片对汞毒害的生理反应常表现在细胞膜透性的增加, 叶绿素、可溶性蛋白和碳水化合物等含量下降, 光合作用和呼吸作用受抑制, 代谢有关酶活性的下降以及自由基累积导致的氧化胁迫增强等。叶绿体、细胞膜等是汞胁迫敏感的细胞器, 而质膜透性、可溶性蛋白和叶绿素含量以及光合作用等常作为汞胁迫下叶片伤害以及抗性比较的适当的生理指标(Patra et al., 2000; 2004; Tripathi et al., 1999)。
已有研究表明稀土元素对刺激植物的生长发育和提高抗性均有一定的作用, 近年来, 应用稀土元素调控植物重金属胁迫的研究已有不少报道, 但有关调控汞胁迫的研究以及以林木为材料的研究尚少, 稀土缓解重金属胁迫的机制有待进一步研究。
多胺与植物重金属胁迫关系密切, 但对于胁迫下植物内源多胺含量的增加是主动的适应性反应还是被动伤害的结果尚存在争议(Shanti et al., 2006; Groppa et al., 2007)。王学等(2003; 2004)研究表明, 外源亚精胺(Spd)可以缓解Hg2+、Cr6+对荇菜(Nymphoides peltatum)的胁迫效应; 刘素纯(2006)的研究认为铅胁迫下腐胺(Put)的累积量可作为黄瓜幼苗抗胁迫效应的生理指标, 是对胁迫的一种适应性反应。尽管已有不少外源多胺和稀土缓解植物重金属胁迫效应的研究报道, 但稀土对重金属胁迫的缓解作用与内源多胺变化之间关系的研究, 目前国内外尚未见报道。
由于重金属污染主要由土壤和水体产生, 有关重金属胁迫的研究多采用联体水培法和盆栽法, 重金属对植物叶片的伤害主要通过对根系的影响所产生的间接作用以及对根系的吸收、运输至叶片积累所产生的直接影响, 在研究重金属对植物叶片的直接影响作用及叶片抗性机制时, 采用离体处理也不失为一种简便、有效模拟方法, 并可消除植物其他部分对叶片的影响。采用离体处理方法的研究报道多见于外文期刊:如Chen等(1999; 2001)用水稻(Oryza sativa)离体叶为材料研究Cu2+胁迫下脯氨酸累积的调节机制以及对抗坏血酸氧化酶的诱导作用; Romanowska等(2002)用离体处理方法研究了Pb2+对大麦(Hordeum vulgare)、玉米(Zea mays)等叶片及其线粒体呼吸作用的影响机制; Ksiazek等(1984)也用离体处理方法研究不同家系杨树叶片对铅的敏感性及其细胞定位。
竹类植物以其用途广、繁殖快、适应力强, 资源丰富, 经济效益、社会效益、生态效益显著而越来越受到林业及园林部门的重视。矢竹(Pseudosasa japonica)为矢竹属的模式竹种, 地下茎复轴混生型, 每节具1分枝, 叶狭长翠绿。原产日本, 为日本著名的园林观赏竹种, 我国江苏、上海、浙江、福建等地的园林引种栽培, 生长良好, 对环境要求一般, 具有较强的抗逆性和观赏性(游为贵, 1998)。本文以矢竹为材料, 采用离体处理方法, 研究镧稀土对汞胁迫下矢竹叶片生理生化反应的调控效应及其与内源多胺变化的关系, 为进一步阐明稀土调节植物叶片重金属胁迫的机制以及园林观赏竹种重金属胁迫生理的研究提供参考。
1 材料与方法 1.1 材料供试竹种矢竹栽植于南京林业大学竹类植物标本园, 于4月份取2年生健壮竹株中层生长状况良好、发育条件一致的成熟叶片用于各种试验处理。
1.2 试验方法 1.2.1 处理方法胁迫处理参照Chen等(2001)的离体处理法。叶片取回清洗干净后去除中脉和两端各1 cm, 剪成约0.5 cm×0.5 cm叶段, 按每份样0.5 g分别置于表 1所示各处理液培养皿中, 处理溶液20 mL, 每日更换1次, 处理温度30 ℃, 每个处理3~6个重复(质膜透性测定6个重复, 其他指标3个重复), 处理时间共8 d。蛋白质、叶绿素取样时间为处理结束后, 其他指标取样时间见图中说明。
|
|
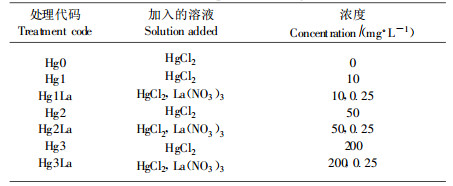
镧稀土为江苏溧阳方正稀土总厂生产的氧化镧(99.9%), 经硝化处理而得, 处理时调pH至6.0, 所采用稀土浓度经预试验确定。
1.2.2 测定方法叶绿素含量测定采用混和(丙酮与无水乙醇的体积比为1:1)浸提法(张志良, 1997); 可溶性蛋白含量的测定采用考马斯亮兰法(Bradford, 1976); SOD及POD活性测定按王爱国等(1983)方法; 内源游离多胺含量采用薄层荧光法测定(谢寅峰等, 1999); 细胞质膜透性测定采用谢田等(1986)的紫外吸收法。每个样品测定3次重复, 数据经Excel及stst统计分析软件处理。
2 结果与分析 2.1 稀土对汞胁迫下矢竹叶片细胞质膜透性的影响以非电解质外渗率表示矢竹叶片细胞质膜透性的变化, 由图 1可知, 不同浓度的Hg2+处理非电解质外渗率在前期(4 d内)均较高, 后期逐渐降低, 表明细胞质膜在前期受到较大程度的伤害, 后期非电解质外渗率的降低可能是由于细胞质膜结构已高度破坏, 细胞内含物前期大量流失所致。从动态变化趋势来看, 不同胁迫浓度表现不同, 从零浓度到高浓度处理非电解质外渗率的高峰分别出现在第5、4、3天和第1天, 表明随着胁迫强度的增加质膜受严重伤害的时间提前、程度加重。低浓度汞胁迫(Hg1)下外源La3+处理总体上降低了非电解质外渗率, 稳定了膜的结构, 尤其在处理前2 d效果明显(图 1a), 而Hg2La和Hg3La处理的非电解质外渗率则分别在第3天和第2天时超过相应的Hg2和Hg3 (图 1b、c), 对膜的伤害起到促进作用, 与Hg1La效果相反。

|
图 1 镧对汞胁迫下矢竹叶片细胞质膜透性的影响 Fig. 1 Effects of La3+ on the plasmalemma permeability in P.japonica leaves under Hg2+ stress |
表 2表明, 不同浓度汞胁迫下矢竹叶片叶绿素含量均呈现明显下降, 胁迫强度愈高, 下降程度愈大。与叶绿素b相比, 胁迫下叶绿素a的下降较为明显, 叶绿素a/b值明显降低。与Hg1相比, Hg1La处理明显抑制了叶绿素a、b及a+b含量的下降, Hg1La比Hg1各叶绿素含量分别增加9.6%、9.1%、9.4%, 差异显著(P < 0.05), Hg2La与Hg2相比虽对各叶绿素含量在胁迫下的下降有一定的抑制作用, 但影响不显著(P > 0.05), 而Hg3La各叶绿素含量则低于Hg3, 且差异显著(P < 0.05), 表明高浓度Hg2+胁迫(Hg3)下La3+处理(Hg3La)对叶绿素含量的下降起促进作用。各浓度汞胁迫下外源La3+处理对叶绿素a/b值影响不明显。
|
|
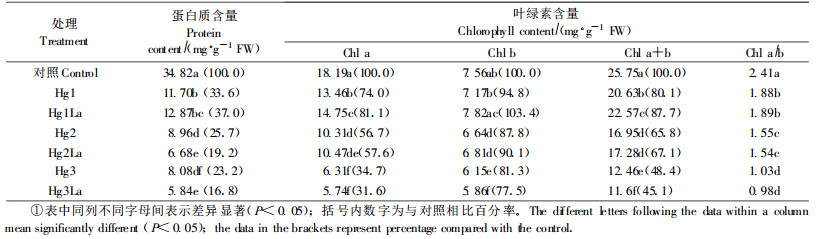
由表 2可知, 矢竹叶片蛋白质含量的变化与叶绿素变化类似。在汞胁迫下蛋白质含量急剧下降, 下降幅度随胁迫强度的增加而增加, Hg1、Hg2、Hg3分别下降66.4%、74.3%、76.8%。与Hg1相比, Hg1La可以减缓胁迫下蛋白质含量的下降, Hg1La比Hg1高10.0%, 而Hg2La、Hg3La与Hg2和Hg3相比则促进蛋白质的降解, 蛋白质含量分别为相应胁迫的74.6%和72.3%, 差异显著(P < 0.05)。
2.4 稀土对汞胁迫下矢竹叶片超氧化物歧化酶(SOD)和过氧化物酶(POD)活性的影响由以上结果可知, La3+对低浓度Hg2+胁迫(Hg1)下矢竹叶片的生理伤害具有缓解作用, 为了进一步探讨缓解作用机制, 对叶片SOD、POD活性以及内源多胺含量的变化进行了比较。
图 2表明:Hg0和Hg1 SOD活性都呈下降趋势, 但Hg1下降明显, 尤其在第1~3天之间, Hg1La处理总体上对胁迫下SOD活性的下降具有缓解作用, 尤其在第5天效果最明显, 比Hg1高63.3%, 第3、8天也分别高于Hg1 8.0%和18.2%。

|
图 2 镧对汞胁迫下矢竹叶 SOD 和 POD 活性的影响 Fig. 2 Effects of La3+ on SOD and POD activity in P.japonica leaves under Hg2+ stress |
POD活性的变化与SOD不同, Hg0在处理过程中基本保持稳定, 而Hg1 POD活性总体都高于处理前水平, 第5天时达到最大值, 为处理前的119.3%。Hg1La处理除第1天超过Hg1外, 其他时间都低于Hg1水平(P < 0.05), 使POD活性基本维持在胁迫前水平, 对胁迫下POD活性的变化具有稳定作用(图 2)。
2.5 稀土对汞胁迫下矢竹叶片多胺含量的影响Hg0和Hg1处理下, 矢竹叶片内源腐胺(Put)都呈现先升后降趋势, 但Hg1增加更为明显和迅速, 1 d后达到胁迫前的1.66倍, 以后有所下降, 但仍高于胁迫前水平。亚精胺(Spd)和精胺(Spm)变化与Put类似, 先升高后下降, 但升高幅度不如Put, 并在胁迫后期低于胁迫前水平, 与Hg0相比Hg1处理后期Spm下降更明显, 多胺总量的变化趋势与Put变化相同。与Hg1相比, Hg1La处理对Put、Spd和Spm的影响趋势相似, 第1天影响不明显; 第3天则使Put、Spd、Spm及多胺总量含量大幅提高, 分别增加48.7%、45.6%、40.0%和47.3%;第5天时增加幅度有所降低, 分别为12.3%、23.9%、9.7%和14.7%, 但差异显著(P < 0.05);到第8天时两者又趋于相近, Put、Spm和多胺总量还略有下降(图 3)。总体而言, Hg1胁迫下, 外源La3+处理(Hg1La)在一定时间内对矢竹叶片内源多胺水平的提高具有显著的促进作用。

|
图 3 镧对汞胁迫下矢竹叶多胺含量的影 Fig. 3 Effects of La3+ on polyamine content in P.japonica leaves under Hg2+ stress |
有关Hg2+、Cr6+、Pb3+等重金属胁迫的许多研究已表明叶片细胞质膜透性、叶绿素和蛋白质含量的变化是反映重金属胁迫条件下植物受伤害程度的重要生理指标。通过对上述指标的综合分析表明, 汞胁迫下矢竹离体叶片受伤害程度与胁迫强度呈正相关。胁迫下叶片叶绿素含量的下降受合成和分解两方面因素的影响, 叶绿素a/b比降低表明Hg对chl a的影响比chl b的影响大, 这与周守标等(2007)通过盆栽试验在其他重金属胁迫下的测定结果一致, 因本文是在离体条件下进行, 因此Hg对chl a的破坏作用比chl b更强可能是主要原因, 其机制尚待进一步研究。另外值得注意的是非电解质外渗率的大小并不能完全代表质膜透性的实际情况, 而应根据其动态变化进行分析, 尤其在胁迫后期非电解质外渗率的降低应是膜透性增大、质膜受伤加重的体现, 这与周长芳等(2001)对水花生(Altemanthera philoxeroides)联体条件下Cu2+胁迫研究中的结果类似。
一定浓度的La3+处理对低浓度汞胁迫下叶片细胞质膜透性的增大、蛋白质和叶绿素含量的降低具有显著抑制作用, 表明对叶片伤害的缓解效应; 中等浓度胁迫下La3+对胁迫前期质膜的伤害具有一定保护作用, 后期则起反作用; 高浓度胁迫下La3+与Hg2+产生协同效应, 加重叶片的生理伤害。庞欣等(2002)通过小麦(Triticum aestivum)联体条件下水培试验的研究也表明了类似的结果。La3+虽然对植物抗Hg2+胁迫有一定的保护作用, 但与胁迫的程度和胁迫时间的长短都具有密切的关系。
稀土对重金属胁迫下质膜结构的保护作用主要表现为2个方面:1)稳定膜结构; 2)影响质子跨膜梯度和膜电位(Zheng et al., 2002)。稀土元素与含氧基团(磷氧键)具有较强的配位能力, 能够直接嵌入质膜中, 取代Ca2+与膜脂大分子形成复杂交联, 或改变膜孔半径与膜分子构型, 使细胞膜结构更趋稳定(Smith et al., 1998); 稀土对膜结构的稳定作用还与其调节活性氧代谢有关, 通过增强胁迫下清除氧自由基能力, 降低膜脂过氧化水平而减轻膜、细胞器及大分子物质破坏, 近年来国内关于稀土缓解重金属胁迫机制的许多研究都与此有关(周青等, 2004)。重金属胁迫下植物体内保护酶活性一般会发生应急性的提高, 以增强清除氧自由基能力, 而本文Hg1胁迫下矢竹叶片SOD活性没有呈现升高现象可能与材料及酶本身特性有关, 与POD相比, SOD应急反应不明显(代全林, 2006), 而对Hg的抑制作用较敏感。另外, 也可能与离体处理方式有关。胁迫下SOD活性持续下降及保护酶系统失调(SOD活性下降, 而POD活性升高)所引起的活性氧清除能力的降低及其代谢紊乱可能是导致矢竹叶片细胞伤害的重要原因, 相对而言, Hg0处理的SOD和POD活性能保持基本稳定, 而Hg1La处理可明显抑制胁迫下SOD活性下降及POD活性升高, 维持保护酶活性的稳定。庞欣等(2002)对Hg2+胁迫、王东红等(2002)对Pb3+胁迫的研究也表明适当浓度的La3+可以提高胁迫下小麦、油菜(Brassica napus)的SOD和CAT活性。但La3+处理下保护酶活性的变化究竟是缓解效应的原因还是结果目前尚不清楚, 从本文POD的表现看似乎只是缓解作用的结果, La3+处理后POD活性的降低是细胞伤害程度减轻的一种表现, 例如La3+也可以直接参与自由基的清除, Wang等(2000)以脉冲辐解法证实, La3+能有效抑制超氧阴离子自由基的生成。但从SOD的变化来分析, 2种可能性都存在, 可能稀土对不同酶所起的调节机制不同, 有关机制尚需进一步研究。
多胺与稀土在调节植物逆境和衰老生理方面具有相似之处, 适当浓度的外施对胁迫伤害都具有一定的缓解作用, 但对于胁迫下稀土调节作用与内源多胺变化关系的研究, 目前国内外尚未见报道。此外, 对于重金属胁迫下植物内源多胺含量的增加是主动的适应性反应还是被动伤害的结果也存在争议。Shanti等(2006)认为重金属胁迫下植物内源多胺含量的增加是积极的适应性反应, 而Groppa等(2007)以向日葵为材料的Cd和Cu胁迫的研究则认为胁迫下Put、Spm的增加是植物受伤害的标志, 尤其在胁迫的中后期。本试验表明:La在缓解Hg1叶片伤害的同时, 进一步提高了内源Put、Spd和Spm的含量, 延缓了胁迫中后期多胺含量的下降趋势, 多胺含量的增加与蛋白质、叶绿素含量以及SOD活性的增加相一致, 而与膜透性及POD活性的变化相反, 推测多胺含量的增高对减轻胁迫下的伤害具有积极的作用, 是对胁迫的一种适应性反应, 而非伤害性反应, 至少La3+的缓解作用与多胺增加有关。王学等(2003; 2004)对外源Spd可以缓解Hg2+、Cr6+对荇菜(Nymphoides peltatum)的胁迫效应的研究以及刘素纯(2006)认为铅胁迫下Put的累计量可作为黄瓜(Cucumis sativus)幼苗抗胁迫效应的生理指标的研究也支持了相同的推论。但由于多胺代谢的复杂性, 不同的植物材料、生理状态、胁迫类型、处理方式等都可能产生不同的结果, 另外, 多胺的作用与稀土一样可能也有浓度效应, 因此, 进一步阐明多胺代谢与重金属胁迫关系还需大量深入的研究。
在逆境下, 多胺与稀土的调节机制有相似之处, 都与稳定膜及大分子物质结构、调节活性氧代谢有关, Shanti等(2006)认为重金属胁迫下多胺主要作为抗氧化剂及“信号分子”参与细胞代谢调节, 多胺含量的升高可能是镧缓解矢竹叶汞胁迫伤害作用的方式之一, 多胺究竟通过何种方式参与La3+对Hg2+胁迫伤害的缓解调节, 还有待进一步研究。
代全林. 2006. 重金属对植物毒害机理的研究进展. 亚热带农业研究, 2(2): 49-53. |
刘素纯.2006.铅对黄瓜幼苗生长发育的影响研究.湖南农业大学博士学位论文
|
母波, 韩善华, 张英慧, 等. 2007. 汞胁迫对植物细胞结构与功能的影响. 中国微生态学杂志, 19(1): 112-113. DOI:10.3969/j.issn.1005-376X.2007.01.052 |
庞欣, 王东红, 邢晓燕, 等. 2002. 汞胁迫下La(NO3)3对小麦幼苗抗氧化酶活性的影响. 中国稀土学报, (4): 159-163. |
王爱国, 罗广华, 邵从本. 1983. 大豆种子的研究. 植物生理学报, 22(1): 77-81. |
王东红, 庞欣, 冯雍, 等. 2002. 铅胁迫下La(NO3)3对油菜抗氧化酶的影响. 环境化学, 21(4): 324-328. DOI:10.3321/j.issn:0254-6108.2002.04.003 |
王学, 施国新, 徐勤松, 等. 2003. 外源亚精胺缓解荇菜Cr6+毒害的生理研究. 环境科学学报, 23(5): 689-693. DOI:10.3321/j.issn:0253-2468.2003.05.024 |
王学, 施国新, 马广岳, 等. 2004. 外源亚精胺对荇菜抗Hg2+胁迫能力的影响. 植物生理与分子生物学学报, 30(1): 69-74. |
谢田, 徐中际. 1986. 测定细胞膜透性的紫外吸收法. 植物生理学通讯, (1): 45-46. |
谢寅峰, 沈惠娟, 李梅枝. 1999. 酸胁迫对林木内源多胺及活性氧代谢的影响. 林业科学, 35(1): 117-121. DOI:10.3321/j.issn:1001-7488.1999.01.019 |
游为贵. 1998. 明溪竹类图志. 南京: 东南大学出版社, 230-233, 171-172.
|
张志良. 1997. 植物生理学实验指导. 北京: 高等教育出版社.
|
周长芳, 吴国荣, 施国新, 等. 2001. 水花生抗氧化系统在抵御Cu2+胁迫中的作用. 植物学报, 43(4): 389-394. DOI:10.3321/j.issn:1672-9072.2001.04.011 |
周青, 黄晓华, 张光生. 2004. 污染生态学领域的稀土应用研究进展. 中国稀土学报, 22(2): 177-182. DOI:10.3321/j.issn:1000-4343.2004.02.002 |
周守标, 王春景, 杨海军, 等. 2007. 菰和菖蒲对重金属的胁迫反应及其富集能力. 生态学报, 27(1): 281-287. DOI:10.3321/j.issn:1000-0933.2007.01.033 |
Bradford M M. 1976. A rapid and sensitive method for the quantation of microgram quantities of protein of protein-dye binding. Anal Biochem, 72: 248-254. DOI:10.1016/0003-2697(76)90527-3 |
Chen Chienteh, Chen Limen, Lin Chuanchi. 2001. Regulation of proline accumulation in detached rice leaves exposed to excess copper. Plant Science, 160(2): 283-290. DOI:10.1016/S0168-9452(00)00393-9 |
Chen Limen, Kao C H. 1999. Effects of irradiance and copper on the activity of ascorbate oxidase in detached rice leaves. Biologia Plantarum, 42(4): 583-588. DOI:10.1023/A:1002631518753 |
Groppa M D, Ianuzzo M P, Tomaro M L, et al. 2007. Polyamine metabolism in sunflower plants under long-term cadmium or copper stress. Amino Acids, 32: 265-275. DOI:10.1007/s00726-006-0343-9 |
Ksiazek M, Wozny A, Siwecki R. 1984. The sensitivity of poplar leaves to lead nitrate and intracellular localization of lead. European Journal of Forest Pathology, 14(2): 113-122. DOI:10.1111/efp.1984.14.issue-2 |
Patra M, Bhowmik N, Bandopadhyay B, et al. 2004. Comparison of mercury, lead and arsenic with respect to genotoxic effects on plant systems and the development of genetic tolerance. Environmental and Experimental Botany, 52(3): 199-223. DOI:10.1016/j.envexpbot.2004.02.009 |
Patra M, Sharma A. 2000. Mercury toxicity in plants. Botanical Review, 66(3): 379-422. DOI:10.1007/BF02868923 |
Romanowska E, Igamberdiev A U, Parys E. 2002. Stimulation of respiration by Pb2+ in detached leaves and mitochondria of C3 and C4 plants. Physiologia Plantarum, 116(2): 148-154. DOI:10.1034/j.1399-3054.2002.1160203.x |
Shanti S, Karl J D. 2006. The significance of amino acids and amino acid-derived molecules in plant responses and adaptation to heavy metal stress. Journal of Experimental Botany, 57(4): 711-726. DOI:10.1093/jxb/erj073 |
Smith J C, Mikiiten T M. 1998. Speciation of yttrium and lanthanides in natural water by inductively coupled plasma mass spectrometry after preconcentration by ultrafiltration and with a chlating resin. Analysis, 123(5): 773-780. |
Tripathi A K, Sadhna T. 1999. Changes in some physiological and biochemical characters in Albizia lebbek as bio-indicators of heavy metal toxicity. Journal of Environmental Biology, 20(2): 93-98. |
Wang Chuenxia, Liu Yali, Li Fengmei, et al. 2000. Inhibition effect of rare earth nitrates on superoxide anion radical by pulse radiolysis. J Rare Earths, 18(3): 286-291. |
Zheng Hailei, Zhang Chuenguang, Zhao Zhongqiu, et al. 2002. Effect of La on H+transmembrane gradient and membrane potential rive seedling roots. J Rare Earths, 20(3): 234-238. |